Pisum Genetics
2008-Volume 40
Research Papers
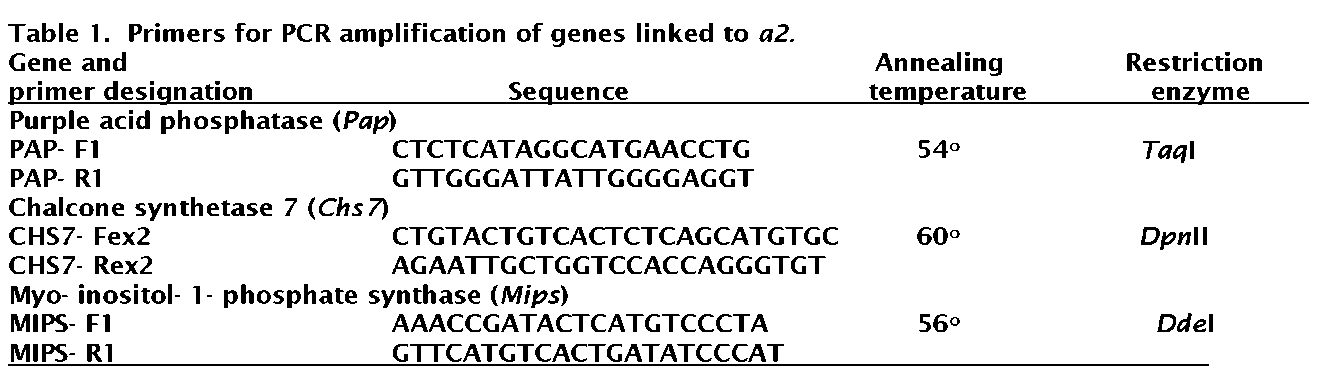
segregating morphological markers, and DNA was extracted from partially expanded leaflets using the procedure of Torres et al. (10). Primers were designed to complement sequences in either Medicago truncatula or Pisumsativum (Table 1). In order to generate gene-specific primers within the CHS family, sequences within introns were used to design primers. Annealing temperatures for the primer pairs and restriction endonucleases used to generate polymorphism are presented in Table 1.
Segregation of the DNA sequences was followed using the CAPS technique (5), resolving the restricted fragments on 2% agarose gels. Due to the complexity of the population, precise recombination distances were not calculated. Instead a 'relative distance estimate' was used to determine an approximate distance between loci. The loci were arranged in a linear order that minimized total number of recombinants. Based on DNA marker results, regions within each line's LG III that appeared to be heterozygous were identified. Recombination events between adjacent loci could then be counted. Two 'distance units' were assigned for a change from a homozygous genotype of one parent to a homozygous genotype of the other but only one unit for a change from a heterozygous genotype to a homozygous genotype. The 'relative distance estimate' between each of the loci and A2 was determined by summing the number of distance units in that interval and dividing by twice the total number of lines for which both loci had been scored (see notes under Table 2). No attempt was made to account for possible recombination between a heterozygous genotype and a dominant phenotype at adjacent loci. Instead all dominant phenotypes adjacent to heterozygous genotypes were treated as heterozygotes.
Results
The scoring of the segregating morphological traits was straightforward except that the pigmentation of flowers on A2_ plants was occasionally faint. The gene b was fixed in the population, so that all A2_ plants had pink flowers. However, it appeared that another gene was affecting the expression of the pink color of the b phenotype in certain lines. Fortunately, leaf axil pigmentation, conferred by dominant alleles at the D locus, was contributed by both parents, and pigmentation in the leaf axil proved to be a more reliable indicator of the presence of the A2 allele than flower pigmentation. All white-flowered plants were examined for lack of pigmentation in the leaf axils and on the testa of the seeds before classifying them homozygous a2.
The purple acid phosphatase primers generated a principle fragment of about 900 bp and several smaller, fainter staining fragments. Restricting the amplified product with TaqI endonuclease produced a multi-fragment pattern indicative of more than a single gene product. TaqI digestion of the 900 bp fragment in Marx line W6 15182 completely digested the original fragment, giving 5 smaller fragments that had a combined size of over twice the original fragment. A similar digest of the original fragment from the a2 parent gave 4 fragments, including the original 900 bp fragment and 3 others that matched 3 of the 5 observed in the digest of the original fragment from the Marx line. I tried several other primers for this gene, all of which gave digests indicating a multigene cluster, and several of which gave different restriction patterns between the two parents. Those polymorphisms that were mapped all co-segregated, suggesting that all polymorphic copies of the gene were clustered.
2
