Biomass production and seed yield of six dry pea cultivars in direct seeding and conventional tillage conditions
Dry pea cultivar development in the region has traditionally been based on conventional tillage practices and their response to direct seeding is unknown. As direct seeding practices gain wider adoption it is necessary to understand the response of pea cultivars to the conditions presented by this production system. In this paper we examine the response of six cultivars grown under conventional and direct seeding conditions over two years. These six cultivars were part of a larger experiment designed to evaluate the U.S. core collection of Pisum germplasm for adaptation to zero tillage (McPhee and Muehlbauer, 2001). The specific objectives of the analysis were to: 1) identify cultivars adapted to direct seeding and 2) quantify genotype by environment interactions.
The six pea genotypes used in this study were ‘Alaska 81’, ‘Joel’, ‘Latah’, ‘Radley’, ‘Rex’ and ‘Shawnee’. The trials were grown at Pullman, WA, on the Washington State University Spillman Research Farm and on a cooperator’s field near Palouse, WA in 1996 and 1997. The Pullman site (46°43’ N, 117°11’ W) was conventionally tilled while the Palouse site (46° 32’ N, 117° 2’ W) had been managed using direct seeding technology for 25 years.
Conventional tillage at the Pullman site involved plowing in the fall following spring barley and spring cultivation followed by sowing with a customized double-disc drill. Seedbed preparation at the Palouse site involved sowing directly into spring wheat stubble with a customized double disc drill. Straw at the Palouse site was harrowed after sowing to facilitate harvest. Cropping sequence at the Pullman location was a spring barley-spring pea rotation and at Palouse a three year rotation of winter wheat–spring wheat–lentil was used. Soil types at the two locations were the Palouse Series: fine-silty, mixed mesic Pachic Ultic Haploxerolls.
Each genotype was replicated seven times and randomly spaced throughout a larger experiment in a completely randomized design both years. Each plot consisted of two rows 1.5 meters long, spaced 0.3 m apart. A single border row of ‘Dark Skin Perfection’ was planted on each side of the plot to reduce border effects. Twenty-five seeds were sown per row resulting in a plant density of 55 plants/m2. Isolated instances of hair pinning were observed at the Palouse site due to heavy cereal residue.
Soil moisture, temperature and precipitation were recorded throughout the growing season (Table 1). Soil moisture was recorded at sowing, flowering and maturity at depths of 0-15, 16-30, 31-45, 46-60 and 61-75 cm at three sites within the plot area (Table 1). Individual plots were harvested when all the plants were beyond physiological maturity. Total aboveground biomass (TAB), including leaves, stems, pods and seeds, was hand-harvested and placed in large paper bags to air dry until all the plots reached moisture equilibrium (approx. 8% moisture). The weight of TAB for each sample was recorded and the samples threshed using a large rubber-roller Vogel plot thresher. Seed yield for each plot was then recorded and the amount of residue was calculated based on the difference between the TAB weight and the seed weight. Harvest index (HI) values for each plot were calculated by dividing the seed yield by the TAB weight.
Total aboveground biomass production at Palouse in 1996 and 1997 was 1850 and 7130 kg/ha, respectively, and 3420 and 6950 kg/ha, respectively, at Pullman. Mean seed yield at Palouse in 1996 and 1997 was 800 and 2930 kg/ha, respectively, and 1490 and 3170 kg/ha, respectively, at Pullman. Mean straw production at Palouse in 1996 and 1997 was 1050 and 4200 kg/ha, respectively, and 1930 and 3780 kg/ha, respectively, at Pullman. The superior total biomass production at Palouse in 1997 was likely due to the greater soil moisture late in the growing season which allowed the plants to continue to produce vegetative matter during the reproductive period and delayed the onset of terminal drought (Table 1). The lower seed yield at Palouse in 1997 compared to Pullman was due to a reduction in seed weight.
Table 1. Summary of sowing date, precipitation received during the growing season, accumulated growing degree days, and total soil moisture at sowing, 50% flowering and maturity at two sites near Pullman and Palouse, Washington in 1996 and 1997.
|
|
|
Precipitation2 (mm) |
|
GDD (°C) |
|
Soil Moisture (%)3 |
||||||||
|
Site1 |
SowingDate |
Veg. |
Repr. |
Total |
Veg. |
Repr. |
Total |
Sowing |
Flowering |
Maturity |
||||
|
1996 |
|
|
|
|
|
|
|
|
|
|
||||
|
Pullman |
May 08 |
100 |
07 |
107 |
449 |
473 |
0922 |
19 |
17 |
14 |
||||
|
Palouse |
May 26 |
015 |
07 |
022 |
553 |
506 |
1059 |
19 |
17 |
15 |
||||
|
1997 |
|
|
|
|
|
|
|
|
|
|
||||
|
Pullman |
May 6 |
045 |
58 |
103 |
566 |
479 |
1045 |
20 |
19 |
18 |
||||
|
Palouse |
May 7 |
054 |
49 |
103 |
599 |
518 |
1117 |
23 |
22 |
20 |
||||
1The Pullman site was managed according to conventional tillage practices and the Palouse site was managed using direct seeding practices.
2Veg. = the vegetative growth period between sowing and flowering; Repr. = the reproductive period from flowering to maturity.
3Percent soil moisture = the average soil moisture between 0 and 75 cm.
Substantial differences in production between 1996 and 1997 were likely due to three factors; 1) total precipitation received during the growing period, 2) distribution of that precipitation and 3) the amount of soil moisture available to the crop. Total precipitation received during the growing season at Palouse in 1996 was 22mm, a reduction of 80 percent compared to Pullman the same year, and only 7mm was received during the reproductive period. The reduced total precipitation during the growing period at Palouse and the marginal amount received during the reproductive period at both locations likely had a negative impact on seed yield and overall plant growth. In comparison, precipitation in 1997 was not substantially different than 1996, but it was distributed more evenly between the vegetative and reproductive growing periods allowing greater plant growth and yield production.
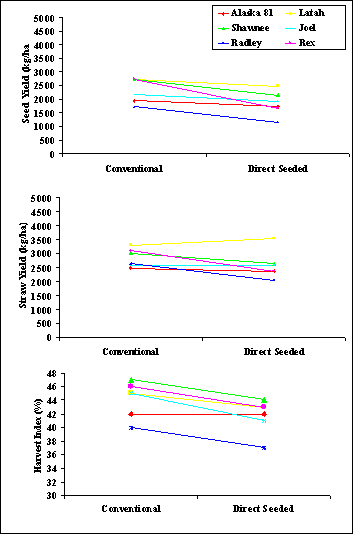
Although analysis of variance showed significant genotype x environment interaction for TAB, seed yield, residue production and harvest index, Rex was the only genotype to show a significant change in rank for all variables and Radley showed a crossover interaction only for straw production (Fig. 1). Therefore, the data generally suggest that little benefit would be realized from additional breeding effort focused on early generation selection in direct seeding environments. Those genotypes which produce well in one set of environmental conditions appear to produce well under other environmental conditions with the exception of environments with specific constraints, such as alkalinity, acute toxic or limiting levels of nutrients, etc. Evaluation of advanced selections under alternative environments should be sufficient to identify those exceptional genotypes with significant advantage in a given environment.
The overbearing constraints to pea crop production reported in the literature are temperature and the amount and distribution of precipitation. These same environmental factors were observed to have great influence in the current study as well. Distribution and amount of precipitation during the growing season appear to have been the primary factors affecting production potential of the genotypes in the specific environments used in this study. Additional research looking at a more diverse collection of genotypes in side-by-side environmental studies may be necessary to identify specific plant characters which can be targeted in selection.
Table 2. Mean production of total aboveground biomass, seed yield, straw yield and harvest index at Pullman (conventional tillage) and Palouse (direct seeded) in 1996 and 1997.
|
Location |
Year |
Total Biomass |
Seed Yield |
Straw Yield |
Harvest Index |
|
(kg/ha) |
(kg/ha) |
(kg/ha) |
(%) |
||
|
Pullman |
1996 |
3417 ± 258 |
1490 ± 142 |
1926 ± 128 |
43 ± 1.4 |
|
|
1997 |
6954 ± 449 |
3174 ± 250 |
3780 ± 219 |
45 ± 1.2 |
|
|
|
|
|
|
|
|
Palouse |
1996 |
1852 ± 250 |
803 ± 129 |
1049 ± 123 |
42 ± 1.2 |
|
|
1997 |
7191 ± 654 |
2960 ± 310 |
4230 ± 372 |
41 ± 1.5 |
Fig. 1. Seed and straw yield for Alaska 81,
Joel, Latah, Radley, Rex and Shawnee grown at Pullman
and Palouse, WA in 1996 and 1997.
1. McPhee, K.E. and Muehlbauer, F.J. 2001. Gen. Res. Crop Evol., 48: 195-203.
2. Muehlbauer, F.J. 1996. Am. J. Altern. Agric., 11: 71-76.
3. Papendick, R.I., Young, D.L., McCool, D.K. and Krauss, H.A. 1985. In: Follett, R.F. and Stewart, B.A. (eds.). Soil Erosion and Crop Productivity. ASA-CSSA-SSA Press, pp 305-320.