Supplemental data on fasciata genes in Pisum
resources
Święcicki W.K.
Inst. of Plant Genetics, Polish Academy of Sciences, Poznań, Poland
Fasciation
is one of the most exciting characters in Pisum.
It was described for the first time in 1597 (3), with different names having
been used for the phenotype by various individuals (Pisum umbellatum, Mummy pea, Crown pea, Pois turc, Pois couronne,
var. coronatum).
Breeders have been interested in possibility of modifying stem
architecture, flowering physiology and maturation using this source of genetic
variation. Unfortunately, the theoretical advantages (most of the pods being
produced on the upper portion of the plant) had associated disadvantages
(lodging and drought stress). Thus,
despite the availability of high yielding fasciata genotypes (5) few cultivars have been released (e.g. Buława/POL,
Ornamenta, Rosacrone and Golf/GER, Novella/USA).
The fasciata phenotype appears to be much more important for
geneticists. It was one of the seven Mendelian characters, and other cases of
spontaneous mutation have been identified.
The use of mutagens have led to the production of a considerable number
of additional mutants (1, 5). Marx
and Hagedorn (9) reviewed the literature dealing with the anatomy, morphology,
expression and inheritance of fasciation in pea. Gottschalk induced and
discribed a similar mutation type: dichotomous
branching. The fasciation in
this mutant type involved only a few nodes, resulting in a forked stem (1, 5).
Gottshalk indicated that the gene bif1
was present in mutants 1201A and 239CH and that bif2
(polymeric to bif1) was present in the
mutant 157A. Both genes showed incomplete penetrance.
Gottshalk (4) also described a mutant, 37B, which proved to be an allele
of the gene bif1 with full penetrance.
Lamprecht
(7) proposed that the fasciata
mutation was controlled by two recessive genes (polymeric and duplicate): fa (12) and fas (6).
The former has been mapped on linkage group IV (6) and the latter on
linkage group III (2). WL 6 (Wt 10006 in the Wiatrowo Genebank) is the type line for
both genes. In contrast, Marx and
Hagedorn (9) concluded that the fasciata
phenotype is controlled by one recessive gene but exhibits variable expressivity
and incomplete penetrance. In the original test cross at Wiatrowo (WL 6 x Wt
3527 and reciprocal) the following F2 segregations were found – 79 Fa
: 20 fa, c2 = 1.21, 77
Fa : 18 fa, c2 = 1.56). Sidorova (11)
tested for allelism among independent fasciata
mutations (induced by different mutagens) and stated that at least 2-3 loci,
supplemented by several modifiers, control this character. Rod and Vagnerova
(10) suggested that the fasciata
phenotype is controlled by three multiple alleles.
Allelism tests were also made by Loennig (8) among 12 fasciata
lines including WL 6. His
results indicated that two of Vasileva’s mutants, Mut I/74 and Mut II/87, were
allelic but differed from the type line at one locus.
The symbol used for the postulated new locus was Fa2.
After
studying many fasciata mutants,
Gottschalk concluded that three to four independent genes with multiple alleles
are responsible for the character (5). Gottschalk stated that “These
interpretations are not necessarily in contradiction to each other...
certain observations indicate that the fasciated pea mutants are not a
uniform group. They are heterogeneous in their morphology as well as in their
genetic constitution.” These
analyses also included the line WL 5544 (weakly fasciated and bifurcated with
unstable penetrance) but did not investigate relationships between fasciata and dichotomous
branching.
The
above, contrasting opinions regarding the genetic basis of the fasciata
phenotype are rather frustrating. The
character could be a suitable gene marker (particularly because linkage group IV
is very poor in markers), but the genotype of many of the fasciata
lines in genebanks is unknown. Moreover,
the only tester line for linkage group IV, WL 1143, has a very weak fasciata expression. In
order to clarify the genetic basis of faciation in pea lines held at the
Wiatrowo Genebank, I performed complementation tests among them in all possible
combinations.
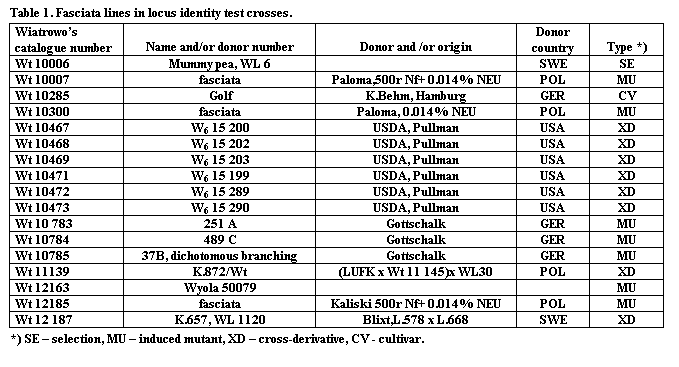
Seventeen lines were identified as possessing a fasciated or similar
phenotype (Table 1). These lines
included material from Blixt, Gottshalk and Marx.
The extensive nature of the full diallele test (272 combinations)
required that the crossings and examination of the F1 plants be
conducted over a period of six years (1996 through 2001) in field at Wiatrowo. The F1 plants, excluding those involving Wt 12 185
and Wt010 785, were all fasciated. The type
line for fa, WL 6, was included among
the 17 lines, indicating that in all lines except Wt 12 185 and Wt 10 785 the
fasciated phenotype is controlled by the fa
gene. In cross combinations with Wt 12 185 the F1 plants were normal
suggesting that the second fasciata
gene from a different locus is involved. When
Wt 10 785 was used as one parent and fa lines as the second the F1 plants displayed the dichotomous
branching phenotype. This result suggests
that the dichotomous branching
mutation is caused by an allele at Fa
with the following dominance (Fa – fabif
– fa).
 There remain several aspects of the genetics of the faciated phenotype
still to be investigated. The
second fasciata gene (in Wt 12 185) needs to be localized on the pea
linkage map. Further allelism tests
need to be conducted including mutant lines from Sidorova, Loennig, and Rod and
Vagnerova. Finally, the
relationship of the bif2 mutations to fabif
and other fasciated mutations needs to be examined.
However, it appears that a tester line for linkage group IV can now be
constructed using an fa gene with
clear expression together with other markers on that linkage group.
There remain several aspects of the genetics of the faciated phenotype
still to be investigated. The
second fasciata gene (in Wt 12 185) needs to be localized on the pea
linkage map. Further allelism tests
need to be conducted including mutant lines from Sidorova, Loennig, and Rod and
Vagnerova. Finally, the
relationship of the bif2 mutations to fabif
and other fasciated mutations needs to be examined.
However, it appears that a tester line for linkage group IV can now be
constructed using an fa gene with
clear expression together with other markers on that linkage group.
0
1. Blixt, S.
1972. Agri Hort. Genet., 30:
1-293.
02. Blixt, S.
1976. Agri Hort. Genet., 34:
83-87.
03. Darbishire,
A.D. 1911. Breeding and the Mendelian Discovery. Cassel and Co., Ltd.,
New York.
04. Gottschalk, W. 1979.
Pisum Newslet., 11: 5.
05. Gottschalk, W.
and Wolff. G., 1983.
Induced Mutations in Plant Breeding. Springer Verlag. p. 238.
06. Lamprecht, H.
1952. Agri Hort. Genet., 10: 158-168.
07. Lamprecht, H.
1974. Monographie der Gattung Pisum.
Steiermärkische Landesdruckerei, Graz. p. 655.
08. Loennig, W.E.
1983. Pisum Newslett., 15: 38-39.
09. Marx, G.A.and
Hagedorn, D.J. 1962.
J. Hered., 53: 31-43.
10. Rod,
J. and Vagnerova, V. 1970. Acta
Univ.Agric. Brne, Fac. Agron., Rada A, 18: 9-15.
11. Sidorova,
K.K. 1970. Genetica USSR, 6: 23-35.
12.
White, O.E. 1917.
Proc. Am. Phil. Soc. 56: 487-589.