New mutations in the linkage group V
have provided stable trisomic lines of pea
Berdnikov,
V.A., Gorel’, F. L.,
Institute of Cytology and Genetics of Russian Academy of Sciences
and
Kosterin, O. E.
Acad. Lavrentiev ave. 10, Novosibirsk 630090 Russia
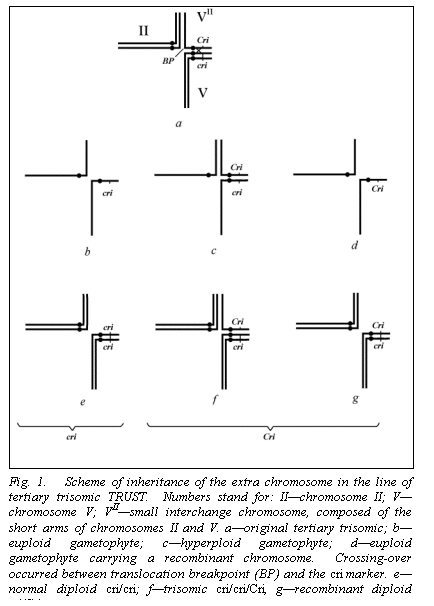 As early as 1939 it was shown that selfing of a heterozygote for the
Hammarlund translocation produced trisomics at a relatively high frequency (up
to 18%) (4). The trisomic karyotype could be represented by seven pairs of
chromosomes of the standard complement plus a small interchange chromosome
composed of the short arms of chromosomes “II” and “V.” (In this paper
we need to refer to chromosomes and their markers; thus we will use the linkage
group numbers for designation of the corresponding chromosomes).
Although viability of these trisomics is lower than that of their diploid
counterparts, they are quite fertile and are easily bred.
In 1992 we made the cross WL1238 x SGE182 (1).
From the F2 generation a plant with a cri,
gp phenotype was selected and crossed
with the line WL1476 bearing the Hammarlund trans-location.
After an insuing test-cross (No. 3 in ref. 1), with a cri,
gp plant as a tester, we obtained a trisomic TRIS-54 in which both
chromo-somes V carried the recessive allele cri,
while the extra chromosome possessed the wild type allele Cri in the vicinity of the breakpoint (BP) (Fig. 1).
As early as 1939 it was shown that selfing of a heterozygote for the
Hammarlund translocation produced trisomics at a relatively high frequency (up
to 18%) (4). The trisomic karyotype could be represented by seven pairs of
chromosomes of the standard complement plus a small interchange chromosome
composed of the short arms of chromosomes “II” and “V.” (In this paper
we need to refer to chromosomes and their markers; thus we will use the linkage
group numbers for designation of the corresponding chromosomes).
Although viability of these trisomics is lower than that of their diploid
counterparts, they are quite fertile and are easily bred.
In 1992 we made the cross WL1238 x SGE182 (1).
From the F2 generation a plant with a cri,
gp phenotype was selected and crossed
with the line WL1476 bearing the Hammarlund trans-location.
After an insuing test-cross (No. 3 in ref. 1), with a cri,
gp plant as a tester, we obtained a trisomic TRIS-54 in which both
chromo-somes V carried the recessive allele cri,
while the extra chromosome possessed the wild type allele Cri in the vicinity of the breakpoint (BP) (Fig. 1).
Among the progeny generated from TRIS-54 we could easily distinguish
diploid plants (genotype cri/cri) from trisomics
(genotype cri/cri/Cri) even as seedlings
(the symbols of genes residing on the extra chromosome are given last.
Additional evidence for the presence of the extra chromosome included
enlarged bracts, larger average number of flowers on inflorescences, and
undulating margins of stipulae and leaflets.
In problematic cases a microscopic analysis of bivalents
in PMC was performed. The
proportion of tisomics in progenies of trisomics varied between 10-20%, so that
selfing permitted a considerable increase in the number of trisomic lines.
In an overwhelming majority of cases, diploid plants resulting from
selfing of trisomics had the phenotype cri,
but among 1052 diploids obtained from trisomic descendants of TRIS-54 we
recorded four recombinant plants with the phenotype Cri.
Thus, the fraction of normal chromosomes V that recombined with the
additional interchange chromosome between the locus Cri and BP was 0.19%.
The recombinant plants differed from trisomics by a greater vigor and
fertility, the bracts were missing and stipulae and leaflets lacked the
characteristic waviness. The
proportion of cri plants in the progeny of the recombinant plants was
approximately 25%.
To introduce additional markers into the short arm of chromosome V we
crossed a trisomic plant (with phenotype u),
resulting from two generations of selfing of TRIS-54, with a diploid plant with
the genotype cri Ust/cri Ust,
originating from the above mentioned cross WL1238 x SGE182 (1).
From the resulting F2 a trisomic with the phenotype Ust
was selected. This plant was the
founder of the near-isogenic trisomic line ‘TRUST.’
This line was increased for several generations, with selection of
trisomics in each until several tens of thousands of plants had been produced.
The proportion of trisomics in each generations of this line was
relatively high (20-30%).
The most striking peculiarity of the line TRUST was an absence of
diploids with the phenotype Cri, indicating suppression of recombination between
Cri and BP of the Hammarlund
translocation. A present we have
scored more than 10,000 individulas and have not found a single recombinant.
The phenotype of TRUST was closer to normal than the trisomic descendants
of TRIS-54: the leaflet margins
were less wavy and pollen fertility and number of seeds per pod were greater.
TRUST trisomics have less than 10% sterile pollen grains as compared to
10-20% in TRIS-54. Nevertheless, in addition to seven bivalents a small
univalent chromosome was invariably observed in PMC.
In reciprocal crosses of the line TRUST with the testerline YELLOW CRISPA
we determined that the extra chromosome was transferred via the female
gametophyte at a frequency of 25.5% (sample size N = 383) and via the male
gametophyte with a frequenacy of 12.3% (N = 983).
We attempted to introduce into the genome of the line TRUST additional
recessive mutations that would be manifested only in diploid plants with the
phenotype cri and covered by normal
alleles in trisomics, with the phenotype Cri.
Approximately 2800 seeds produced by TRUST trisomics were left to imbibe
in 0.1% EMS solution. At the
seedling stage plants with the phenotype cri
were removed and the remaining 789 plants grown in a greenhouse.
For the majority of plants, seed productivity appeared to be very low.
The seeds collected from the 210 most productive M1 plants were sowed in
the greenhouse. From each family a
single M2 trisomic plant exhibiting no visible mutations was chosen,
and the progenies of such plants were planted in the field.
By selecting the plants with the normal phenotype we tried to eliminate,
at least in part, induced mutations not covered by the extra chromosome.
Among 207 M3 families 43 displayed various mutations but only three of
these exhibited a strong linkage with cri.
One, called fas3, had a
fasciated stem, while two other, called killer
(kil) and remover (rmv) were
believed to be recessive lethals because no plants of the phenotype cri
were observed in the families where they were found.
The trisomic carries of the mutations rmv
and kil became founders of the lines TRUST-R and TRUST-K, respectively.
The line TRUST-K does not differe from the initial line TRUST, either by
the phenotype of trisomics, by pollen fertility, or by number of seeds per pod.
However, after imbibition only about 25% of the seeds germinate to give
rise to trisomics, while the germination process of the remaining 75% of the
seeds stops when the radicals are about 5 mm long.
A screening of 1012 seedlings of the line TRUST-K with the genotype cri
kil/cri kil/Cri Kil revealed
only one diploid plant with the phenotype cri.
Thus, the fraction of the normal chromosome V recombining with the extra
chromosome between Kil and Cri was 0.05%.
Two plants of the line TRUST-K were crossed with the line WHAF (crdwh,
a, af) described in ref. 2. Two
resulting F1 plants wre diploids with the phenotype (A,
Cri). Of 154 F2 plants examined, only six had the
phenotype cri, indicating that the location of the sporophytic lethal kil
is on the short arm of chromosome V
about 6.0 + 1.4 cM from the locus Cri
(computed by the maximum likelihood method using equations from ref. 3).
Thus, in trisomics the recombination distance between the loci Cri
and Kil is reduced at least 100-fold.
The line TRUST-R is remarkable in that all its seeds germinate and give
rise only to trisomics cri rmv/cri rmv/ Cri
Rmv. The number of seeds per pod in this lines is reduced by half.
Most probably, the diploids die at an early stage of embryo development.
Screening of more than a thousand plants of this line failed to identify
a single plant with the phenotype cri,
giving a recombination distance between Cri
and Rmv of less than 0.05 cM.
A plant of the line TRUST-R was crossed with the line WHAF.
An F1 plant was trisomic with the genotype
cri rmv/Cri Rmv/Cri Rmv. In this plant
recombination between the chromosomes of the basic complement could easily
occur, but among 60 F2 plants none with the phenotype cri
was found. It is evident that the lethal rmv
is tightly linked to cri.
In addition, we obtained three other lines for which the yield of
trisomics had changed significantly. In
two of them, TRUST-6 and TRUST-138, the proportion of trisomics was about 50%,
wihle in the line TRUST-111 the fraction was 90%.
At present we are investigating the nature of the mutations underlying
this increase in the proportion of trisomics.
In summary, based on the Hammarlund translocation we have created a
genetic system allowing easily differentiation between diploids and trisomics
for a short interchange chromosome with little effect on viability and
fertility. Due to an unknown event
that occurred between TRIS-54 and TRUST, the ability of this extra chromosome to
recombine with those of the normal karyotype has been sharply reduced.
Finally, through the use of chemical mutagenesis we induced two lethals
that exclude diploids from the progeny, in effect producing two stable trisomic
lines.
1. Berdnikov,
V.A. Gorel’, F.L. and Temnykh, S.V. 1993.
Pisum Genetics 25: 18-21.
2. Berdnikov,
V.A. and Gorel’, F.L. 2000.
Pisum Genetics 32: 6-8.
3. Kosterin,
O.E., Berdnikov, V.A. and Gorel’, F.L. 1999.
Pisum Genetics 31: 33.
4. Sutton,
E. 1939. J. Genet. 38: 459-476.