|
Pisum Genetics |
Volume 26 |
1994 |
Research Reports |
pages 34-35 |
Recombination within the complex locus His(2-6) containing genes for five histone H1 subtypes in pea
|
Trusov, Y.A., Bogdanova, V.S. and Berdnikov, V.A. |
Institute of Cytology and Genetics Novosibirsk 630090, Russia |
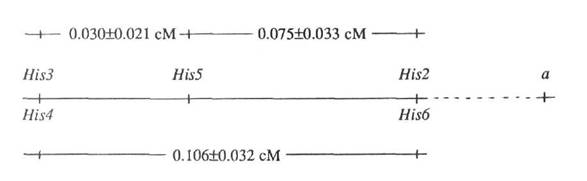
It is now reliably established that the pea genome contains at least 7 genes that encode histone H1 proteins. They are mapped to three loci: His1 in linkage group V (4), and His(2-6) and His7 in linkage group I (1, 3). Locus His(2-6) consists of 5 closely clustered genes situated within 4.1±1.5 cM of the anthocyanin gene, a (1).
This paper describes an attempt to register recombination events between the genes of locus His(2-6) and to determine its fine structure. Each gene of this locus has allelic variants. Their protein products are easily identified by electrophoresis in 15% polyacrylamide gel containing 8M urea/0.9M acetic acid. Isolation of histone H1 is described in (3). Following Berdnikov et al (2), the H1 allelic combination of a plant (the 'haplotype') is designated by a formula which in this case includes five positions, because it denotes the allelic states of five genes (His2 to His6) encoding H1 subtypes 2 to 6. In this formula, the numerals denote allelic forms of the genes and their positions in the formula correspond to the gene number (the first position to His2, the second to His3, and so on). The allelic forms were numbered in accordance with their relative electrophoretic mobility. Among four variants of subtype 5 there are two with close electrophoretic mobility which were not resolved by Berdnikov et al (2). These two variants were denoted as 2' (the slow form) and 2 (the fast form).
The experiment involved more than 3000 individual F2 plants from four independent crosses (see Table 1). Parental lines with particular haplotypes were obtained from the VIR collection: VIR6560 (haplotype 21133) ; VIR4871 (22122) ; VIR5195 (11123) ; VIR5417 (2212'1) ; cultivar Torsdag (21211) ; a plant with haplotype 11223 was obtained from the F2 progeny of cross VIR5195 (11123) with Sprint-1 (21221) (an extra rapid line derived in our laboratory from the cross Avanti x VIR7036).
The recombinants observed in these crosses are listed in Table 1. Taking into account the scarcity of recombinants, we combined data presented in the table for all crosses and mapped individual genes of the locus His(2-6) as follows:
We did not find recombinants between His3 and His4 or between His2 and His6. For this reason, while combining the data from different crosses, we treated these genes pairs as single units, although in several cases only one gene of a pair had different alleles in parental forms.
The parental lines in cross 2 differed from each other also in alleles of the anthocyanin gene, a (the plants with 11223 had allele A and those with 2212'1 had allele a). The genotype for the anthocyanin locus was determined only in recombinants by their progeny. The genotype at the a locus occurred in parental combination with the corresponding alleles of His2 and His6 in all cases, but never with alleles of His3 and His4. The result suggests that this histone cluster is oriented to the gene a by its His2 His6 end.
Table. 1 Genotypes of F2 plants with recombinant haplotypes for the His2-6 gene cluster.
|
|
Cross 1 |
Cross 2 |
Cross 3 |
Cross 4 |
|
Parents |
21133 x 21211 |
11223 x 2212'1 |
11123 x 21211 |
22122 x 21211 |
|
Number of F2 |
266 |
1716 |
763 |
568 |
|
Genotypes of |
none |
11223/1212'3 |
11123/21111 |
22122/21212 |
The recombinant haplotypes are printed in boldface
The data obtained in this study can shed light on an intriguing fact which was observed in extensive investigation of peas cultivated all over the Old World (1). No plant was found with the haplotype 22XX (the haplotypic formula refers only to genes His3 to His6 because gene His2 was represented chiefly by allele 2; X means any allele), while haplotypes 12XX and 21XX were found to be abundant in the same regions. At the same time, the most common alleles of gene pairs other than His3 and His4 were found in all possible combinations. This might be explained in two ways: 1) very tight linkage and 2) some functional prohibition. The supposition that recombination was involved in formation of the diversity of haplotypes, being extremely rare or even impossible between His3 and His4 genes, remains only a first explanation.
Our map could be helpful while designing new combinations of histone H1 genes. It is very interesting to construct and examine a plant with haplotype 20000 (zero means that protein products of the corresponding genes are absent). We have crossed lines with 20101 and 21020 and have already obtained the combination 20100.
Belyaev, A.I. and Berdnikov, V.A. 1981. Genetika (USSR) 17:498-504.
Berdnikov, V.A., Bogdanova, V.S., Rozov,
S.M. and Kosterin, O.E. 1993. Heredity
71:199-209.
Kosterin, O.E. 1992. Pisum Genetics 24:56-59.
Rozov, S.M., Bogdanova, V.S. and Berdnikov,
V.A. 1986. Genetika (USSR)
22:2159-2166.