|
Pisum Genetics |
Volume 23 |
1991 |
Research Reports |
pages 29-34 |
Internode length in Pisum: le5839 is a less severe allele than Mendel's le
|
Ross, J.J. and J.B. Reid
|
Department of Plant Science, University of Tasmania Hobart, Tasmania 7001, Australia |
Mutant alleles at the le locus, particularly Mendel's le (5, 16), have proved valuable in unravelling the role of gibberellins (GAs) in the control of internode length in Pisum sativum (1, 13). The allele le partially blocks the conversion of GA20 to GA1 the bioactive gibberellin in peas (1, 2), and consequently confers the dwarf phenotype. Quantification of GA1 levels in the apical portions of isogenic LeLe and lele lines (using an internal standard and gas chromatography-mass spectrometry) has shown that tall (Le Le) plants typically contain 10-18 times more GA1 than comparable dwarf plants (14). However, similar determinations on another isogenic pair of lines, Torsdag (tall, LeLe) and NGB5839 (dwarf, le5839le5839) yielded a somewhat smaller difference in GA1 level (5-6 fold; 11). This suggests that the mutant allele in NGB5839, le5839 (an induced mutation, 3) may be "leakier" than Mendel's le. However, this is not supported by the very short internode length of NGB5839 (3). Here we examine this question further at the phenotypic level. The evidence comes from a linkage study in which a gene pair linked to the le locus (V, normal pods / v, sugar pods) was used to monitor the inheritance of the mutant allele present in NGB5839. The v and le loci are linked (10) with an overall RCV of 12.6 ± 0.47% (4). The effect of a photoperiod extension with incandescent light on internode length in lele and le5839le5839 plants is also examined.
Materials and Methods
The pure lines used were Nordic Gene Bank line 5839 (NGB5839) (VV le5839le5839), NGB463 (vv lele) and cv. Dippes Gelbe Viktoria (VV lele). NGB5839 and Dippes Gelbe Viktoria carry allele Lf (minimum flowering node 11) while NGB463 carries lfa (minimum flowering node 5; 6). NGB5839 was produced by mutagenesis from cv. Torsdag, by Dr K.K. Sidorova (Novosibirsk, U.S.S.R.).
The plants were grown in a heated glasshouse. The day temperature was usually 20-25°C and the night temperature was 15-18°C. The growing medium was a 1:1 mixture of dolerite chips and vermiculite, topped with 3-4 cm of potting mix. For generations F1 to F6 the light regime consisted of natural daylight extended with mixed fluorescent (Thorn 40 W cool white tubes) and incandescent (Mazda 100 W pearl globes) light (intensity ca. 30 mmol m-2 s-l at pot top) to give a photoperiod of 18 h. Certain F6 plants were grown in either an 8 h photoperiod (8 h natural light) or a 24 h photoperiod (8 h natural light extended to 24 h with weak incandescent light at an intensity of ca. 3 mmol m-2 s-l at pot top). The flowering node is defined as the number of the node bearing the first initiated flower, counting from the cotyledons as zero.
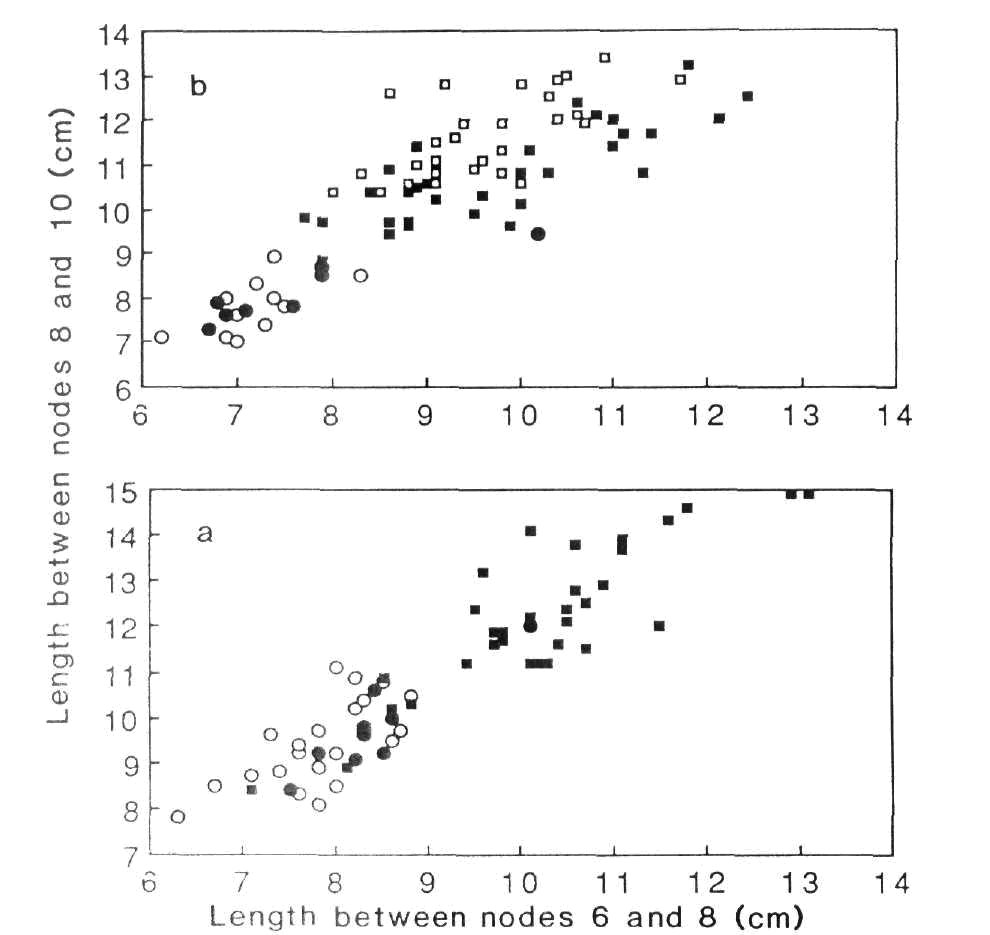
Fig. 1. Stem length between nodes 8 and 10 plotted against stem length between nodes 6 and 8 for F5 (a) and F6 (b) plants from cross NGB463 x NGB5839. Key: ○, vv plants from vv parents; ●, vv plants from Vv parents; ■, V - plants from Vv parents; □, VV plants from VV parents. The F5 was generated from 4 vv and 3 Vv F4 plants, and the F6 from 2 vv, 3 Vv and 2 VV F5 plants.
Results
The F1 of cross NGB463 x NGB5839 was dwarf, as were all plants in subsequent generations. Generations F3 to F6 were produced by single plant selection beginning with an F2 segregate of genotype lfalfa Vv. Plants of genotype vv (sugar pod) possessed, on average, significantly (P < 0.001) shorter internodes than V- (normal pod) plants in all generations from F2 to F6 (e.g. Table 1, Fig. l, a and b). This suggests that the mutant allele in NGB5839, le5839 (which entered the cross linked to V), results in longer internodes and is, therefore, a leakier allele than le.
Table 1. Stem length (cm) between nodes 6 and 9 for lines NGB463 and NGB5839, and for Lf- V- and Lf- vv segregates in the F2 generation of crosses NGB463 x NGB5839 and NGB463 x Dippes Gelbe Viktoria. The data are shown as mean ± SE with n in parentheses. Photoperiod 18 h.
|
Line or Cross |
Genotype |
Stem length |
|
NGB463 |
lfalfa vv |
16.10 ± 0.50 ( 6) |
|
NGB5839 |
LfLf VV |
6.32 ± 0.29 (6) |
|
NGB463 x NGB5839 F2 |
Lf- V- |
10.41 ± 0.35 (50) |
|
|
Lf- vv |
7.50 ± 0.35 (11) |
|
NGB463 x Dippes Gelbe Viktoria F2 |
Lf-V- |
16.33 ± 0.46 (43) |
|
Lf- vv |
16.18 ± 1.28 (10) |
Table 2. Stem length (cm) between nodes 6 and 9 for le5839le5839 and lele segregates from cross NGB463 x NGB5839 grown in either an 8 h or a 24 h photoperiod. Data are shown as mean ± SE of 10 replicates.
|
Genotype |
Stem length |
|
|
8 h photoperiod |
24 h photoperiod |
|
|
le5839le5839 |
11.91 ± 0.30 |
18.01 ± 0.56 |
|
lele |
8.14 ± 0.17 |
10.97 ± 0.24 |
While there was some overlap of internode length values between the presumed lele and le5839- plants in earlier generations, this was minimal by F6 (Fig. 1, b). From F2 to F6, short plants of genotype vv produced only short offspring (e.g. Fig. 1, a and b). Plants of genotype V- were usually taller, while the vast majority (e.g. 89% in F5 and 88% in F6) of vv segregates from Vv parents were short. However, several taller vv segregates from Vv parents were observed; in one such case from the F4, the progeny was grown and comprised 5 short and 12 taller types (data not shown). This suggests that this F4 plant was of genotype vv le5839le (a recombinant). The F5 generation included 5 short V- segregates (Fig. 1, a). The F6 generation from one of these plants was grown and consisted of 1 short vv plant and 13 taller V- plants. Thus this F5 plant was clearly not a recombinant; the reason for its short stature is not clear. However, it is noteworthy that all VV F6 plants possessed considerably longer internodes than all vv F6 plants from vv parents (Fig. 1, b).
In contrast to the results from cross NGB463 (vv lele) x NGB5839 (VV le5839le5839) segregation of the gene pair V/v was not associated with differences in internode length in cross NGB463 (vv lele) x Dippes Gelbe Viktoria (VV lele) (Table 1).
Although the number of individuals available for comparison was small, the internode length of heterozygous le5839le plants was intermediate between that of homozygous le5839le5839 and lele plants. For example, in F3 the mean values for the stem length between nodes 6 and 9 for le5839le5839, le5839le and lele plants were (in cm) 17.4 ± 1.40 (n=3), 14.43 ± 0.92 (n=4) and 11.67 ± 0.03 (n=3), respectively. It therefore appears there is very little dominance of either allele over the other.
When le5839le5839 and lele plants with a similar genetic background (F6 segregates descended from a single F4 plant from cross NGB463 x NGB5839) were grown in 8 h and 24 h photoperiods, the internodes of both genotypes were longer in 24 h (8 h natural light plus 16 h incandescent light) than in 8 h (Table 2), in accordance with previous results (e.g. 8, 11). However, the response shown by le5839le5839 plants (a 51 % increase) was greater than that of lele plants (a 35% increase, Table 2). There was no evidence that this difference was due to factors other than the genotype at the le locus. For example, the flowering behaviour of both le5839le5839 and lele plants was similar. Both groups initiated flowers at nodes 6-8 in both photoperiods and in the 24 h photoperiod flower development ensued either at the node of initiation or at one node higher. In the 8 h photoperiod substantial flower abortion occurred, but to a similar extent in both genotypes. (The flowering genotype of these plants therefore appears to be lfalfa EE SnSn DneDne, see 7).
Discussion
In cross NGB463 (vv lele) x NGB5839 (VV le5839le5839), V le58399- segregates, on average, possessed longer internodes than did vv lele segregates. In NGB5839, GA1 levels were not reduced to the same extent as in lele lines (compared with isogenic LeLe lines; 11,14).
Table 3. Effect of segregation for the le5839/le pair of alleles on the number of seeds per plant in the F5 and F6 generations of cross NGB463 x NGB5839. The data are shown as mean ± SE with n in parentheses.
|
Generation |
Genotype |
|
|
le5839- |
lele |
|
|
F5 |
28.47 ± 0.78 (30) |
23.55 ± 0.60 (33) |
|
F6 |
18.53 ± 0.36 (59) |
15.63 ± 0.41 (19) |
Considered together, these results strongly suggest that allele le5839 imposes a less severe block on GA1 biosynthesis than does le. Clearly le5839 is a different allele from le and the designation le5839 should remain to indicate this. On the basis of measurements of true-breeding le5839le5839 and lele F6 families (descended from a single F4 plant) allele le5839 increases stem length between nodes 6 and 9 by ca. 40% in an 18 h photoperiod, compared with lele plants. The paradoxial aspect of the present and previous work is that NGB463 (lele) possesses much longer internodes than NGB5839 (le5839le5839) (Table 1). Clearly the two pure lines differ with respect to other loci which affect internode length (e.g. possibly at the Cry locus). The presence of lfa in NGB463 would most likely also result in longer internodes. However, this cannot alone explain the stature of NGB463, since this line was considerably taller than lfalfa lele (and lfalfa le5839-) segregates in F2-F6 (data not shown).
The existence of at least one proven recombinant (genotype le5839 le vv; phenotype long internodes, sugar pods) in cross NGB463 x NGB5839 strongly indicates that the short stature of vv plants in F2 to F6 of this cross is not due to a pleiotropic effect of v. This is confirmed by the lack of effect on internode length of segregation for V/v in cross NGB463 x Dippes Gelbe Viktoria, which, furthermore, is consistent with the presumption that both NGB463 and Dippes Gelbe Viktoria possess the "normal" le allele.
The identification of allele le5839 increases to four the number of alleles at the le locus (in order of increasing length, led, le, le5839 and Le, see 12). It seems possible that allele le5839 may be of some agronomic value since it has the effect of increasing internode length compared with le. In this context it is of interest that in the F5 and F6 generations of cross NGB463 x NGB5839 (on a lfalfa EE SnSn DneDne genetic background in an 18 h photoperiod, see 9), le5839- plants produced ca. 20% more seeds than did lele plants (P < 0.001; Table 3). However, this effect cannot for certain be attributed to the difference at the le locus since the plants also differed at the v locus.
It is well known that in pea internode elongation is enhanced by photoperiod extensions with incandescent or far-red-rich light (3, 8, 11, 15). It has also been shown (3, 11) that line NGB5839 (le5839le5839) responds to such extensions to a lesser extent than does its tall (LeLe) progenitor, Torsdag. In contrast, some le dwarf lines or selections are at least as responsive as Torsdag (Table 1 from 8; Torsdag = L107). However, in the present study le5839le5839 plants responded to an incandescent photoperiod extension to a greater extent than did lele plants (Table 2). Therefore the difference in responsiveness between NGB5839 and le lines referred to above is probably attributable to differences in genetic background. The results shown in Table 2 support the suggestion (3) that on a constant genetic background the response to an incandescent (or far-red-rich) photoperiod extension decreases as the severity of the genetic block in GA1 biosynthesis increases (see also 11).
Acknowledgement. We thank the Australian Research Council for financial support.
Ingram, T.J., J.B. Reid, I.C. Murfet, P.
Gaskin, C.L. Willis and J. MacMillan.
1984. Planta 160:455-463.
Ingram, T.J., J.B. Reid and J. MacMillan. 1986. Planta 168:414-420.
Jolly, C.J., J.B. Reid and J.J. Ross. 1987. Physiol. Plant. 69:489-498.
Lamprecht, H. 1948. Agri Hort. Genet. 6:10-48.
Mendel, G. 1866. Verhand. Natürf. Ver.
Brunn 4:3-47. (Reprinted 1951 in J. of
Hered. 42:3-47).
Murfet, I.C. 1975. Heredity 35:85-98.
Murfet, I.C. 1985. In Handbook of
Flowering, Vol. IV. Ed. A.H. Halevy,
CRC Press, Boca Raton, pp. 97-126.
Murfet, I.C. 1988. Ann. Bot. 61:331-345.
Murfet, I.C. 1990. PNL 22:78-86.
Rasmusson, J. 1927. Hereditas 10:1-150.
Reid, J.B., O. Hasan and JJ. Ross. 1990 J Plant Physiol. 137:46-52.
Ross, JJ. and J.B. Reid. 1987. Ann. Boi. 59:107-109.
Ross, J.J., J.B. Reid, P. Gaskin and J.
MacMillan. 1989. Physiol. Plant.
76:173-176.
Ross, J.J., J.B. Reid and U.S. Dungey. 1991. Planta (in press).
Wellensiek, S. J. 1973. Scientia Hort. 1:77-83.
White, O.E. 1917. Proc. Amer. Phil. Soc. 56:487-589.