38
PNL Volume 21 1989 RESEARCH
REPORTS
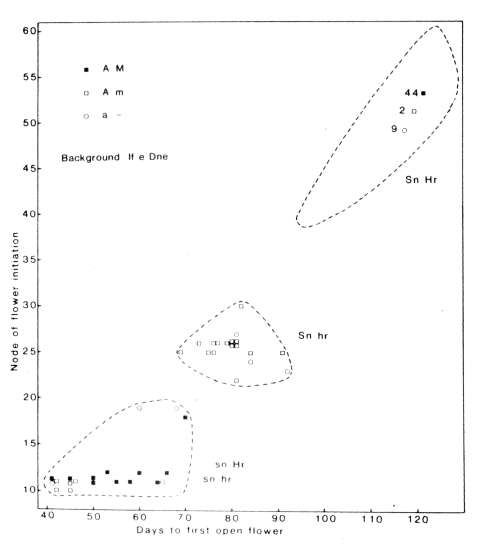
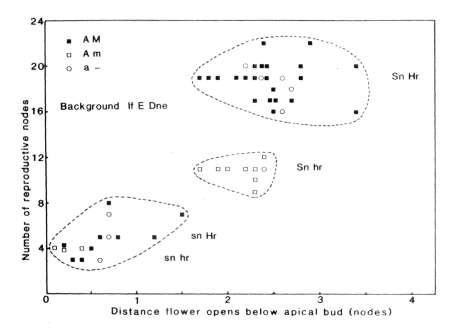
1. Node of
flower initiation and days to
first open flower for F2
plants of
cross 64 (A lf e sn Dne Hr M) x 53 (a lf e Sn Dne hr m) grown in an 8 h photoperiod; day
temperature about 23 C, night 15-18 C. The genotypes for the flowering
genes Sn-sn and Hr-hr, flower colour (red A, white a) and seed
marbling (present M, absent m)
are indicated in the figure.