PNL Volume
20
1988 RESEARCH
REPORTS
15
1. Straub, J. 1940. Ber. Dtsch. Bot. Gas.
58:430-436.
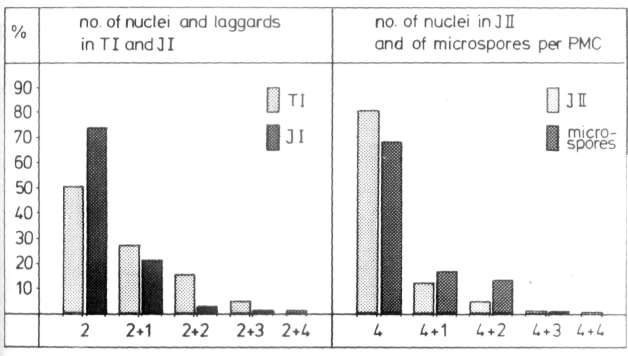
Fig. 1.
The meiotic irregularities of the tetraploid Pisum mutant 408B.
|
|
||
|
14
PNL Volume 20
1988 RESEARCH
REPORTS |
||
|
|
||
|
TETRAPL0IDY FOLLOWING X-IRRADIATION
Gottschalk, W.
Institute of Genetics,
University of Bonn
Federal Republic of Germany
Very rarely, polyploid mutants arise through
mutagens which usually induce gene and chromosome mutation but not genome
mutations. Such an event occurred in our X-ray treatments in
Pisum sativum after seed irradiation with 9 kr. In a very
small M2 family, a plant with reduced fertility was found. It produced
only three seeds from which three tetra-ploid M3 plants developed (mutant
408B of our collection). They were morphologically normal but their seed
production was considerably lower than expected for "normal" tetraploid
individuals. One of them was studied cytologically in order to determine
the causes of the strongly reduced fertility.
Unfortunately, the garden pea is
cytologically a very unfavorable species. Only a very limited number of pollen mother cells (PMC) was
available and it was not possible to analyze the meiotic behavior during
the first prophase and metaphase. From the final stages of the first
division, however, it could be concluded that, besides
bivalents, quadriva-lents had been present in the earlier stages and that the reduction
of chiasma frequency had led to univalents in relatively high
frequencies. They appeared as laggards in the final stages of the first
division. In telophase I, one to four Laggards per PMC were found in almost 50% of all PMCs
studied (Fig. 1, left). Between telophase I and interkinesis, about half
of the lagging chromosomes
were included into the two
daughter nuclei and the proportion of PMCs with micronuclei was correspondingly
lower. It cannot be expected that these processes occurred in a regulated
manner resulting in genomically
balanced nuclei. On the contrary, many of the interkinesis nuclei were
certainly unbalanced with
regard to both chromosome number and genomic constitution.
The proportion of PMCs with disturbances was greater in the
final stages of the second meiotic
division than at the end of the first
division. In 19.2% of all interphase II PMCs studied, one to four separated
chromatids were present outside of the "normal" nuclei. In PMCs already
containing microspores, the proportion of cells of .this category was higher
(31.5%, Fig. 1, right ). It is
not c Lear how these additional anomalies between interphase II and
microspore formation arose. Some of the chromatids, which apparently belonged to distinct nuclei, obviously were not included in these nuclei; on the contrary, they developed independently
into small microspores.
Almost 70% of tho PMCs studied formed four microspores.
From the meiotic irregularities observed, it can be concluded that many of
them were genomically unbalanced, being unable to produce functional germ
cells. This may have been the main reason for the low seed production of the
tetraploid M3 plants. All their seeds were sown in the
following year but exclusively diploid plants developed from them. Thus,
the mutant got lost. Blixt (personal communication) also has observed
cytological instability of the tetraploid level of Pisum
sativum. Tetraploid peas, obtained after colchicine treatment,
lost their tetraploid valence in succeeding generations and their
progenies reverted to the diploid level. Similar experiences were reported
by Straub (1). |
||
|
|
||
|
|
||||
|
PNL Volume
20 |
1988 RESEARCH
REPORTS |
15 |
||
|
|
||||
|
1. Straub, J. 1940. Ber. Dtsch. Bot. Gas.
58:430-436. |
||||
|
|
||||
|
|
||||
|
|
||||
|
Fig. 1.
The meiotic irregularities of the tetraploid Pisum mutant 408B. |
||||
|
|
||||