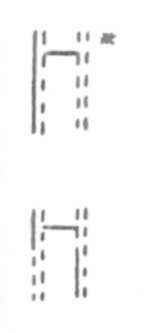86
PNL Volume 19
1987 RESEARCH REPORTS
SISTER CHROMATID EXCHANGE (SCE) IN PISUM
SATIVUM
Wolff, G.,
A. Neumann, and R. Rapp
Institute of Genetics, University
of Bonn
Federal Republic of Germany
Federal Republic of Germany
After application of BrdU
(Bromdesoxyuridine) spontaneous
sister chromatid exchange (SCE) was first demonstrated by Latt (2)
in human chromosomes. This method subsequently was improved, and
SEC was observed in a great number of mammalian chromosomes as
well as in plant cells (Vicia faba [1], Allium cepa [3], and
others). Recently, we could show SCE in Pisum sativum root tip
chromosomes. We applied the method of Kihlman and Kronberg (1)
with a few modifications. Root tips were exposed to BrdU concen-
trations of 10-4 m and 10-6 m in tap water. To increase the
number of metaphases isovalerylurea was used instead of colchi-
cine. The tissue was macerated in a solution of pectinase (5%)
and cellulase (1%). After staining with Hoechst 33258 and Giemsa
the two chromatids of the metaphase chromosomes were differen-
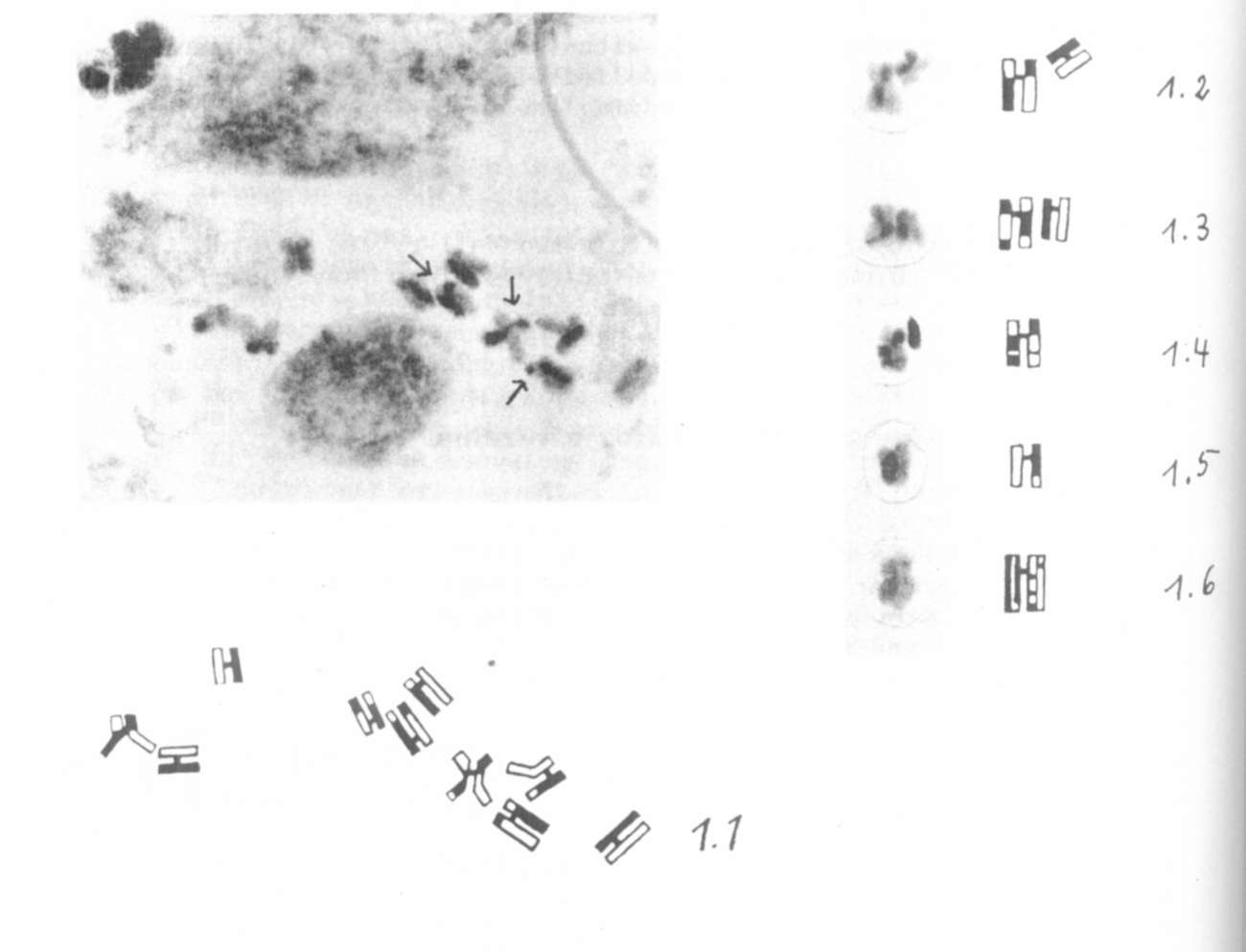
tially stained and SCE's could be detected (Fig. 1.1 and 1.2-1.4,
arrows indicating SCE's). We investigated two series of prepara-
tions :
sister chromatid exchange (SCE) was first demonstrated by Latt (2)
in human chromosomes. This method subsequently was improved, and
SEC was observed in a great number of mammalian chromosomes as
well as in plant cells (Vicia faba [1], Allium cepa [3], and
others). Recently, we could show SCE in Pisum sativum root tip
chromosomes. We applied the method of Kihlman and Kronberg (1)
with a few modifications. Root tips were exposed to BrdU concen-
trations of 10-4 m and 10-6 m in tap water. To increase the
number of metaphases isovalerylurea was used instead of colchi-
cine. The tissue was macerated in a solution of pectinase (5%)
and cellulase (1%). After staining with Hoechst 33258 and Giemsa
the two chromatids of the metaphase chromosomes were differen-
tially stained and SCE's could be detected (Fig. 1.1 and 1.2-1.4,
arrows indicating SCE's). We investigated two series of prepara-
tions :
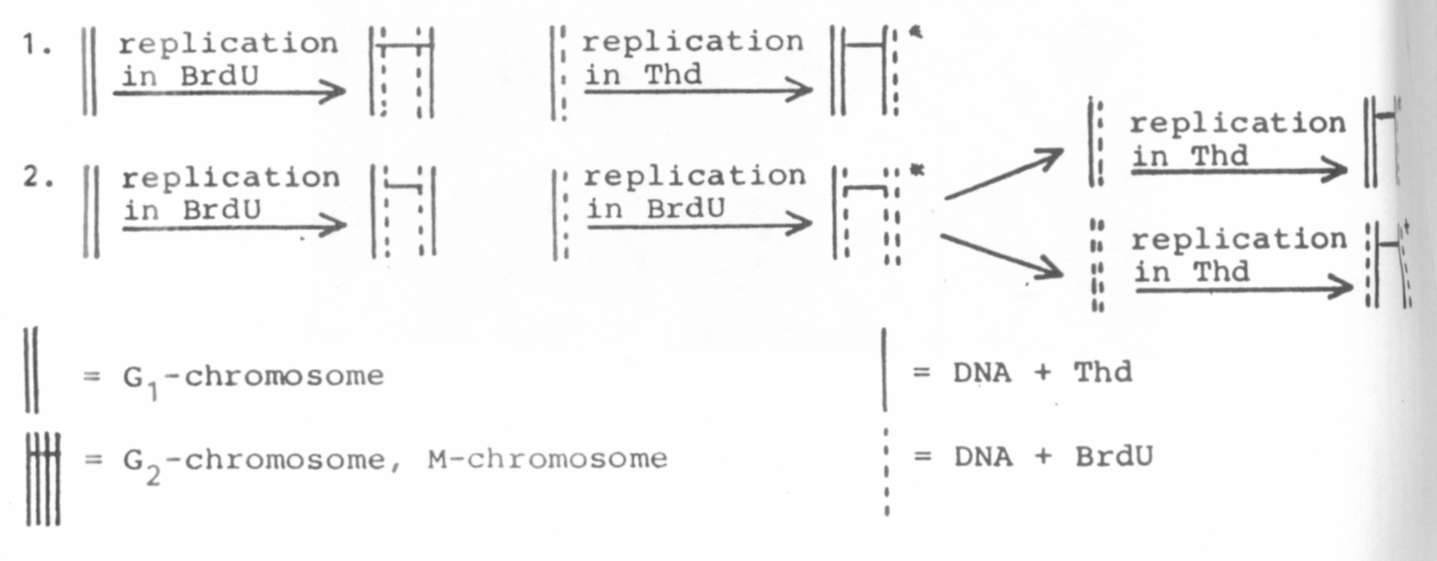
1. Root tips were exposed to BrdU during one
cell cycle,
followed by one cycle in Thd (Thymidine).
followed by one cycle in Thd (Thymidine).
2. Root tips were exposed to BrdU during two
cell cycles,
followed by one cycle in Thd.
followed by one cycle in Thd.
All solutions were made with tap
water, Schematically, the incor-
poration of BrdU results in the following chromosomes:
poration of BrdU results in the following chromosomes: