PNL Volume 19 RESEARCH REPORTS 27
tions or through fluctuations in
the concentration gradient of the
signal; one of the two LTM's gets into the area of concentration
above the threshold and the other into the one beneath it. In
this way, a tendril/leaflet pair might develop at once and at the
same leaf nodium (Fig. 2).
signal; one of the two LTM's gets into the area of concentration
above the threshold and the other into the one beneath it. In
this way, a tendril/leaflet pair might develop at once and at the
same leaf nodium (Fig. 2).
Whether or not the series of
molecular processes, as des-
cribed in the preceding paragraph, will lead to a certain form has
to be shown in a simulation of a diffentiated elaborated model.
An imaginable general starting point for such a model has been
introduced in the first paragraph. However, the translation of
such a starting point into a concrete model and its simulation
involves numerous theoretical and practical problems. In addi-
tion, quite a number of empiric studies, as well as a statistical
recording of the forms of leaves and their deviations (not pro-
duced by mutations), are necessary.
cribed in the preceding paragraph, will lead to a certain form has
to be shown in a simulation of a diffentiated elaborated model.
An imaginable general starting point for such a model has been
introduced in the first paragraph. However, the translation of
such a starting point into a concrete model and its simulation
involves numerous theoretical and practical problems. In addi-
tion, quite a number of empiric studies, as well as a statistical
recording of the forms of leaves and their deviations (not pro-
duced by mutations), are necessary.
1. Ingensiep, H.
W. 1986. PNL 18:67-68.
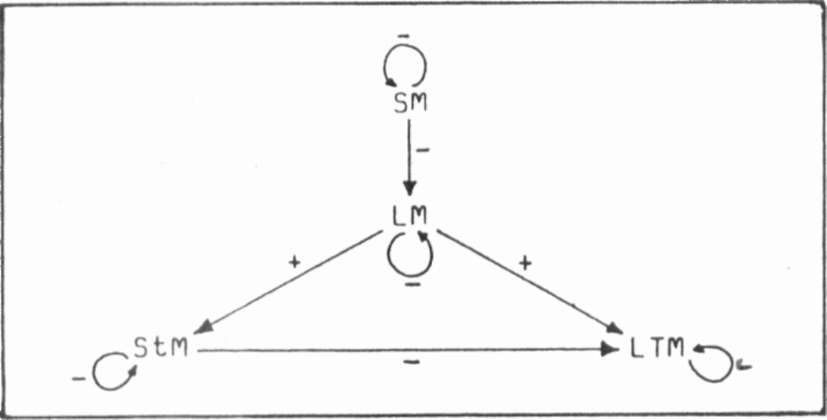
Fig.
1
Interactions among the meristems
during the development
of a leaf of Pisum sativum. + : Inhibition up to a
certain distance, furtherance from this distance on.
- : Inhibition up to a certain distance, no furtherance
from this distance on. (Further explanations in the text.)
of a leaf of Pisum sativum. + : Inhibition up to a
certain distance, furtherance from this distance on.
- : Inhibition up to a certain distance, no furtherance
from this distance on. (Further explanations in the text.)
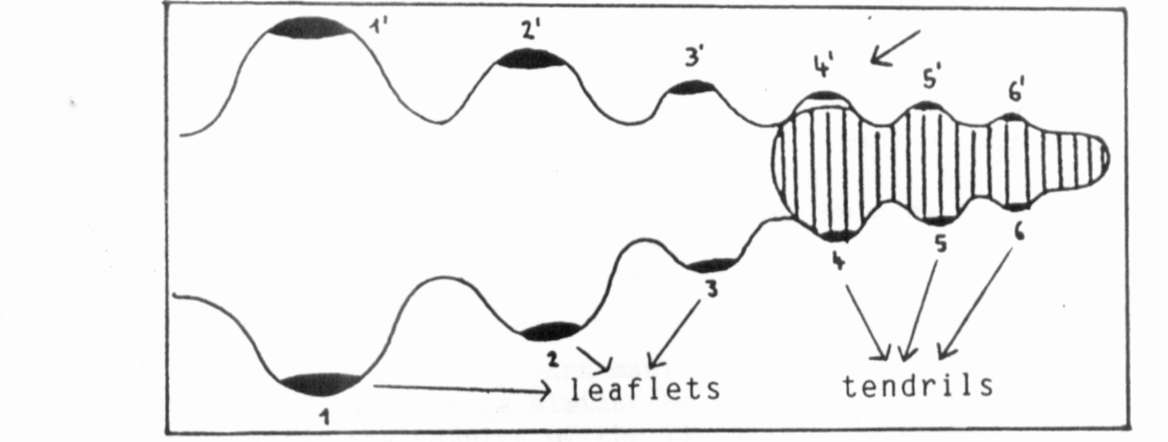
Fig. 2. Explanation of the induction of leaflet-tendril pairs
because
of an asymmetric gradient: 4" is induced to become a leaflet
instead of a tendril.
of an asymmetric gradient: 4" is induced to become a leaflet
instead of a tendril.
*****