PNL Volume 19 1987 RESEARCH REPORTS
at afila and acacia, a form of leaf arises which shows numerous
ramifications and lengthened leaf internodes, as well as leaflets
only. Such a leaf form corresponds to that of the recombinant
afila/acacia. However, the leaflets of this recombinant, compared
with those of the wild-type and those of acacia, respectively, are
very small. The model does not yet account for this and other
characteristics.
ramifications and lengthened leaf internodes, as well as leaflets
only. Such a leaf form corresponds to that of the recombinant
afila/acacia. However, the leaflets of this recombinant, compared
with those of the wild-type and those of acacia, respectively, are
very small. The model does not yet account for this and other
characteristics.
This can only be considered as a first attempt to integrate
the mutations afila and acacia into the phyllomorphogenetic model.
A number of leaf mutations, e.g. apulvinic and cochleata, cannot
be explained on the basis of the hypothetical mechanisms presen-
ted. Perhaps, however, the speculations will supply us with new
starting points for questions of an experimental and theoretical
kind.
the mutations afila and acacia into the phyllomorphogenetic model.
A number of leaf mutations, e.g. apulvinic and cochleata, cannot
be explained on the basis of the hypothetical mechanisms presen-
ted. Perhaps, however, the speculations will supply us with new
starting points for questions of an experimental and theoretical
kind.
1. Ingensiep, H. W. 1986. PNL 18:67-68.
2. Lenz, J. and H. W. Ingensieg. 1987. PNL 19:25.
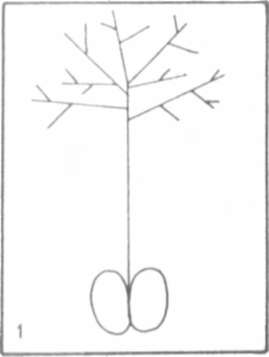
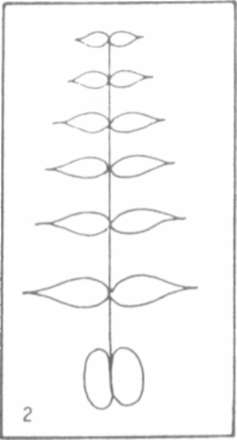
Fig.1.
Schematic of an afila leaf.
Fig. 2. Schematic of an acacia leaf.
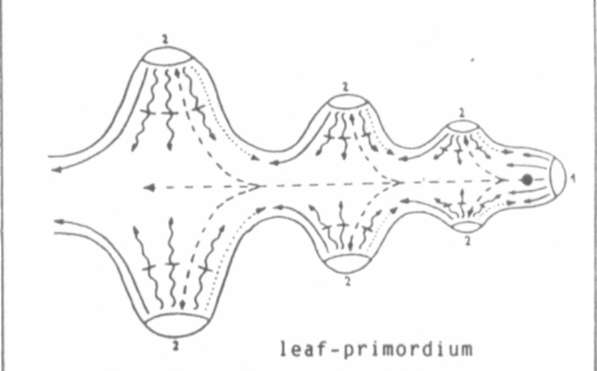
Fig.3
Some basic mechanisms during
phyllomorphogenes is and starting-points for the mutations afila and acacia.
leaf-tip meristem
leaflet/tendril meristem
signal which isolates LTM's
leaflet/tendril meristem
signal which isolates LTM's
signal which promotes the synthesis of the following signal
signal which decides whether leaflets or tendrils are to be formed
auxin
signal which decides whether leaflets or tendrils are to be formed
auxin
starting-point of the mutation afila
starting-point of the mutation acacia
starting-point of the mutation acacia

*****