PNL Volume 18 1986 RESEARCH REPORTS
51
Table 1. Analysis of F2 populations derived from the cross
A M Tac Apu St x A m tac apu st.
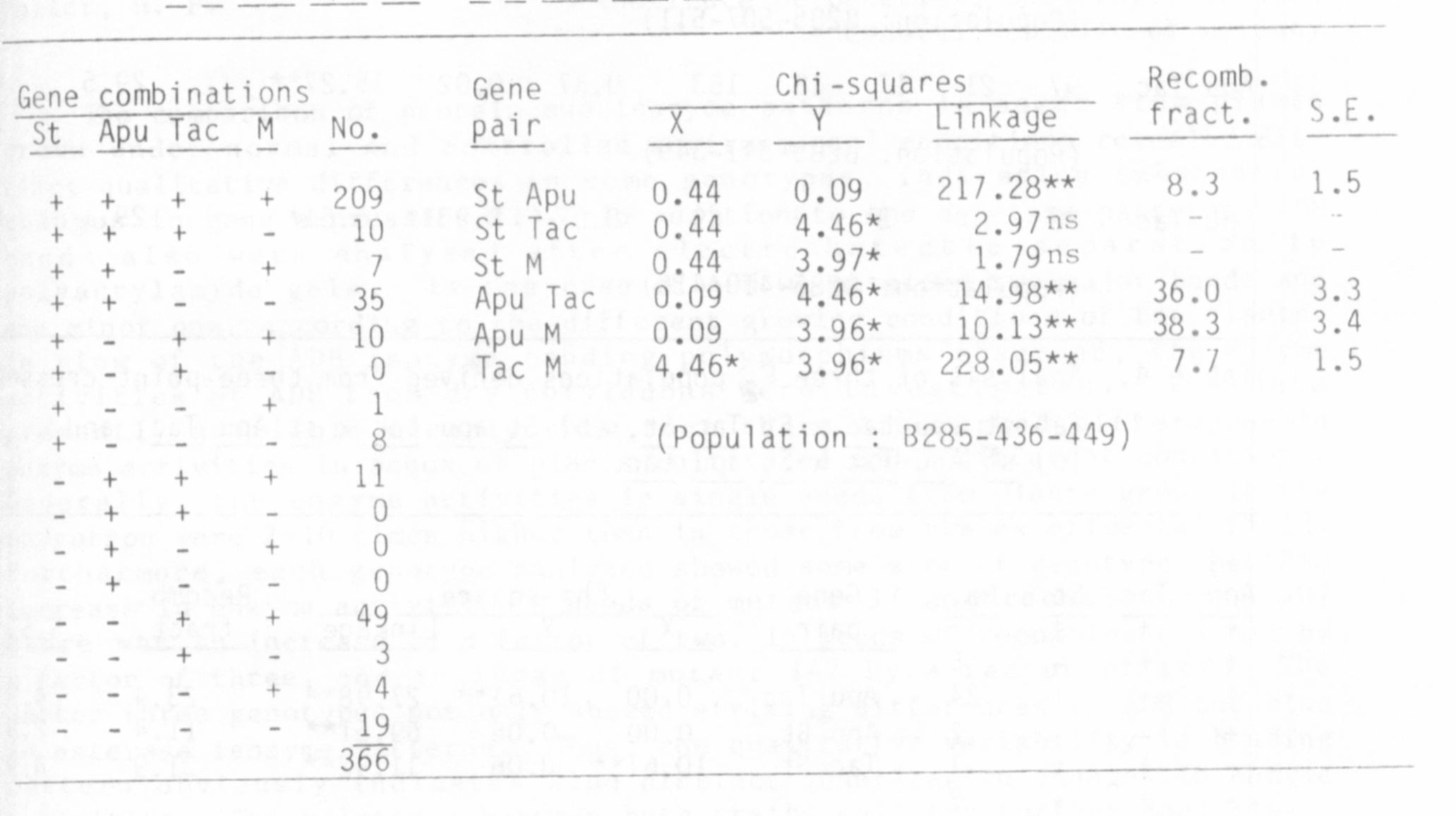
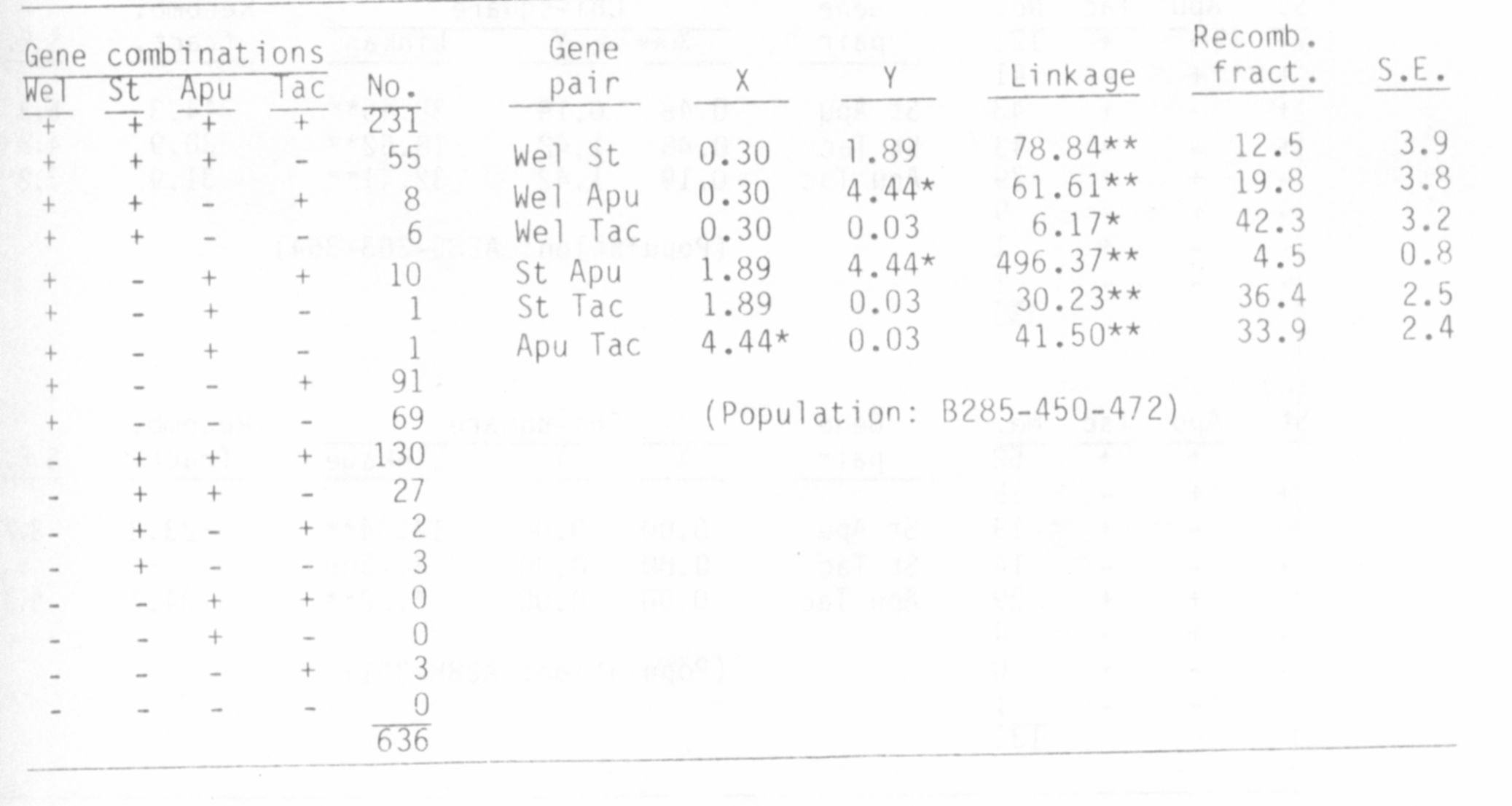
Table 2. Analysis of F2 populations derived from the cross
Tac Apu St wel x tac apu st Wel.
|
|
|||
|
PNL Volume 18 1986 RESEARCH REPORTS
|
49
|
||
|
|
|||
|
TENDRILLED ACACIA (tac): AN ALLELE AT THE Uni LOCUS
Marx, G. A. NYS Agricultural Experiment Station, Geneva, NY USA
Tendrilled acacia (tac) recently has been found to be situated in
chromosome 3 (5), toward the M end of the chromosome (6). To fix its
position more precisely, tac was crossed with lines which were known or
thought to reside in the segment between st and uni. It became apparent
from these crosses that tac and uni are allelic.
Analysis of crosses involving st apu tac and M (Table I) revealed a
rather close linkage between M and tac, and the close association between
st and apu, reported previously (5), was confirmed. Since earlier evidence
(3,4) indicated that we 1 is located close to st, in the region between st
and M, we1 was also used in crosses with tac. The present evidence,
however, indicates that wel lies toward the b end of chromosome 3 rather
than toward the M end. The substantial population size in the present
studies and the fact that the cross is a four point cross lends con-
siderable credence to the new findings.
Tables 3-4 provide additional linkage data for chromosome 3 markers
collected in the course of pursuing studies not directly related to this
investigation.
Since M and uni are known to be closely linked (2) and inasmuch as tac
and M are closely linked (Table 1), tac and uni were expected to be closely
linked as well. To test this assumption, tac was crossed with WL 187 (type
line for uni), seeds of which were kindly supplied by Dr. Blixt. Since
uni/uni plants are sterile, it was necessary to use a number of phenotypi-
cally normal segregants in WL 187 as parents to ensure recovering the uni
allele from a heterozygous plant. A total of 73 F1 seeds from 11
individual F1 plants were planted. Six of the F1 progenies contained wild-
type plants exclusively whereas five progenies contained plants of two
different phenotypes: some wild type and some resembling a combination of
tac and uni. In plants of the latter class the early-formed leaves were
unifoliate; later leaves became tripartite, followed by two pairs of
leaflets and a terminal leaflet. These plants characteristically had no
terminal tendrils, but rarely an odd subterminal tendril did appear.
Flowers on the affected plants were somewhat malformed In contrast to the
flowers borne on sibling plants. The malformation notwithstanding, the
flowers more closely resembled normal flowers than the distinctively
sterile inflorescences of uni/uni plants. Fertility was only somewhat
impaired. The described plants were interpreted as carrying the tac allele
and the uni allele together in the same plant, thus evidencing alleles at a
single locus.
To pursue this supposition further, and to demonstrate that the plants
in question were not selfs, I grew the selfed seed of two of the F1's
showing the described hybrid phenotype. One population contained 44 F2
plants (one died early), the second 18 plants (Table 5). The spectrum of
plant phenotypes was the same in both populations. There were no normal,
wild-type plants in either population. Most plants resembled tac (some
with typical subterminal tendrils) and a minority resembled the distinctive
phenotype conferred by uni/uni, including the typical malformed, sterile
|
|||
|
|
|||
|
|
|||
|
50
|
PNL Volume 18 1986 RESEARCH REPORTS
|
||
|
|
|||
|
inflorescences (Table 6). The tac allele apparently is dominant to the uni
allele but the dominance is incomplete and the heterozygotes usually can be
distinguished from tac homozygotes (and readily from uni homozygotes).
Segregation for these phenotypes was accompanied by normal segregation for
other markers present in the cross (Table 6); segregation also occurred at
the R and I loci.
Even before recognizing the stated allelism certain similarities were
evident between tac and uni plants in the early seedling stage. The first
true leaf of tac plants often is unlfoliate and, conversely, uni typically
bears some tripartite leaves, although later in development.
Another feature of tac plants is the reduced number of leaflet pairs.
In fact, this is a convenient means to distinguish tac tl from Tac tl in
segregating populations. Sharma (7) has already pointed out that the
terminal leaflet of tac plants is not appreciably reduced in size as is
typically the case in _t_l plants. In plants homozygous for tac and for tl,
the pair of subterminal tendrils (characteristic of tac) are absent, thus
for this property tl is epistatic to tac. However, tac Tl plants often
show variability of expression with respect to the subterminal tendrils;
not infrequently the leaves have only one tendril or they may have none.
Thus the absence of subterminal tendrils is by itself not a sure way to
distinguish between tac t1 and Tac t1 plants. Such a distinction can
confidently be made, however, by observing differences in the number of
leaflet pairs per leaf. Plants with the tac t1 combination have fewer
leaflet pairs than those with tl alone; thus, in this respect, tac is
epistatic to tl. In an af_ background the distinction between tac tl and
tac Tl/- is unmistakable. Moreover, the difference between af tac tl and
af Tac tl is evident by the larger and fewer laminae in the former than in
the latter. (This difference evidently was also recognized by Sharma (7)
because, in his diagramatic illustrations, he depicts the leaflet size of
af tac t1 plants as larger than af Tac t1 plants). Thus, tac is an inter-
esting and powerful mutant gene in a number of respects.
There remains the question of symbolization. Because the discovery
of unifoliata (1) antidates tendriled acacia, perhaps the symbolization
should be: Uni,Unitac ,uni. According to Sharma and Kumar (8) there already
exist two alleles of tac: tacb , and tacs . Whether Uni is a classical
multiple allelic locus or a complex locus remains to be seen.
|
|||
|
|
|||
|
1. Lamprecht, H. 1933. Hereditas 28:269-296.
2. Lamprecht, H. 1948. Agri Hort. Genet. 6:10-48.
3. Marx, G. A. 1972. PNL 4:30-31.
4. Marx, 6. A. 1974. PNL 6:30-31.
5. Marx, G. A. 1984. PNL 16:46-48.
6. Marx, G. A., N. F. Weeden, and R. Provvidenti. 1985. PNL 17:57-60.
7. Sharma, B. 1981. Pulse Crops News1 . 1(1):56-57 .
8. Sharma, B. and S. Kumar. Pulse Crops Newsl. l(3):2 1-22.
|
|||
|
|
|||
|
|
|||
|
PNL Volume 18 1986 RESEARCH REPORTS
|
51
|
||
|
|
|||
|
Table 1. Analysis of F2 populations derived from the cross
A M Tac Apu St x A m tac apu st.
|
|||
|
|
|||
|
|
|||
|
Table 2. Analysis of F2 populations derived from the cross
Tac Apu St wel x tac apu st Wel.
|
|||
|
|
|||
|
|
|||
|
|
|||
|
|
||
|
52 PNL Volume 18 1986 RESEARCH REPORTS
|
||
|
|
||
|
Table 3. Analysis of populations derived from three crosses of the
constitution: apu tac x Apu Tac,
|
||
|
|
||
 |
||
|
|
||
|
Table 4. Analysis of three F2 populations derived from three-point crosses
(a) st apu tac x St Tac St, (b) St_ a£u tac x st Apu Tac, and
(c) st Apu Tac x St apu tac.
|
||
|
|
||
 |
||
|
|
||