Marx, G. A. NYS Agricultural Experiment Station, Cornell University
Geneva, NY USA
Plants possessing the EMS-induced recessive mutant curl (6) bear a
clear and distinctive phenotype early in the seedling stage and throughout
their ontogeny. Though curl segregants are weaker and less productive than
Curl/- counterparts, the mutant is a favorable marker for genetic studies.
Preliminary observations made several years ago indicated that curl might
belong to chromosome 7 (data not shown).
In 1984-1985 I constructed populations to verify and refine the
linkage relationship among curl, r, and bt. The results of these crosses,
three in all, are given in Table 1. For one population the F1 seeds were
separated into round (R/-) and wrinkled (r/r) classes before planting but
each F2 plant was also classified for R-r again at the dry seed stage to
ensure the accuracy of the classification.
The data argue persuasively that curl is linked with r and bt in what
is still generally accepted as chromosome 7, the Indicated order being
curl-r-bt. Since, however, there still is controversy concerning the
placement of the r-tl-bt group (2,3) curl is an additional clear marker
which may help to resolve the controversy. Another unsettled matter con-
cerns the placement of wsp on chromosome 7. I have not succeeded in
confirming the reported linkage relationship between wsp and chromosome 7
markers (4). The fact that curl appears to be located toward the wsp end
of chromosome 7 on the current map should aid in the ongoing quest to
settle this issue as well. The linkage relations of two other newly dis-
covered markers may also contribute to a resolution.
Orc, described by Blixt and Swiecicki (1,7), is a dominant mutant that
causes orange cotyledons containing high amounts of lutein (5). My own
experience with Ore indicates that the lower leaves of mutant plants become
orangy yellow about the time the plants begin to flower, thus providing
another diagnostic criterion for scoring Or c plants in segregating
populations. A moderate sized F2 segregating simultaneously for Bt-bt,
Orc-orc, and R-r indicates that the three genes are linked (Table 2).
Accordingly, Orc and curl should show strong linkage, a supposition we are
in the process of testing.
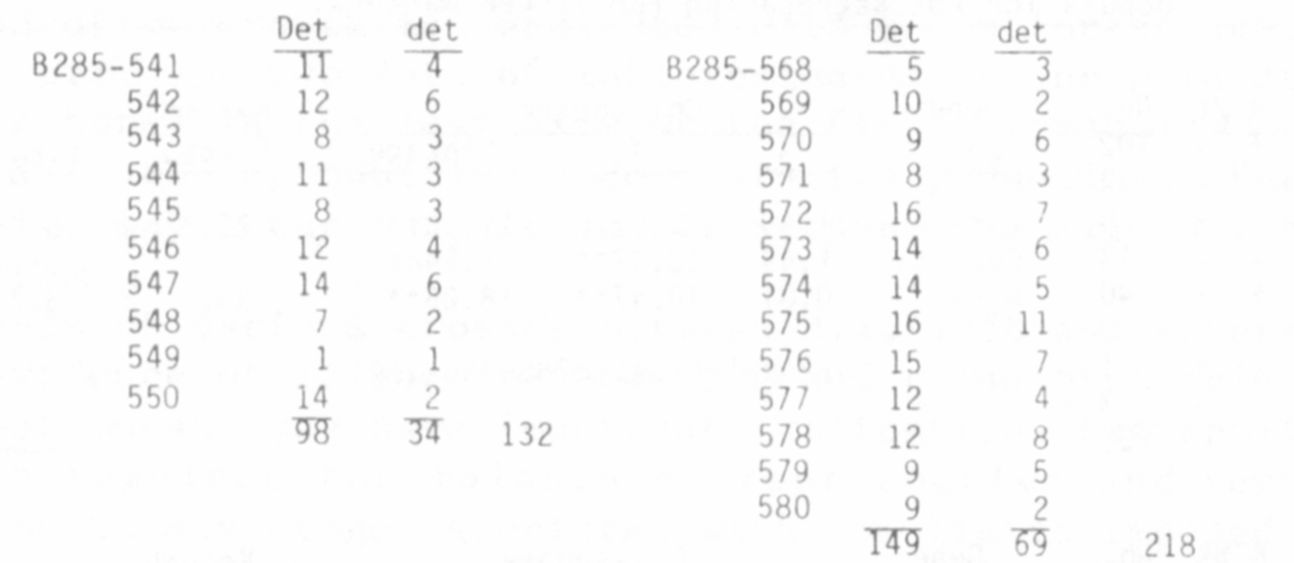
Another mutant that has recently come into my possession, and which I
will provisionally designate det— , also shows linkage with R and Bt. Seed
of the mutant was provided through the courtesy of Dr. Peter Matthews; the
mutant line is maintained in the John Innes Institute Pisum collection
under the number J.I. 1358/B. The mutant behaves as a monogenic recessive
(Table 3a) and is distinguished from normal, wild type plants by its deter-
minate reproductive behavior. The term "determinate" has been used in
1/ It falls to Drs. Matthews and Blixt to accept or reject the gene name and
symbol.