A SIMPLE ASSAY FOR GENOTYPE-DEPENDENT AUXIN-SENSITIVITY WITH YOUNG
SEEDLINGS OF PISUM SATIVUM
Ingensiep, H. W. Institute of Genetics
University of Bonn, Federal Republic of Germany
Based on earlier investigations and considerations about auxin-
dependent morphogenetic response of pea seedlings (1, 2), a simple assay
has been developed for pre-screening auxin-sensitive genotypes in the
young seedling stage using 2,4-D, NAA, and IAA. The idea was to try to
correlate morphogenetic differences (from the initial line) with some
alteration(s) in the endogeneous auxin control system of the plants. The
main regulation points for the latter may be on the level of auxin syn-
thesis, auxin reception or auxin inactivation. The irreversible degrada-
tion of auxin by oxidation and the possible reversible conjugation to
amino acids seem especially important to establish endogenous gradients
of the free auxin controlling shoot eleongation or axillary bud elonga-
tion. Therefore seedlings of the initial line (IL), 'Dippes Gelbe Vik-
toria', were compared with a recombinant (R 650 A) which contains genes
for long internodes, afila, fasciation, and extremely late flowering in
an assay system which may give some hint if these deviations are due to
some alteration in genes involved in the system of auxin inactivation.
Seedlings of IL and R 650 A grown in moist vermiculite for about one
week (light/dark=16/8 hr, 25C, 50% RH) were decapitated 1 cm above the
second node and prepared pieces of chromatography paper,dipped in
ethanolic solutions of the auxins up to 10-2M, were inserted at the top.
After 24 hr the pieces were removed and the seedlings were observed
during the following period of cultivation under the same conditions.
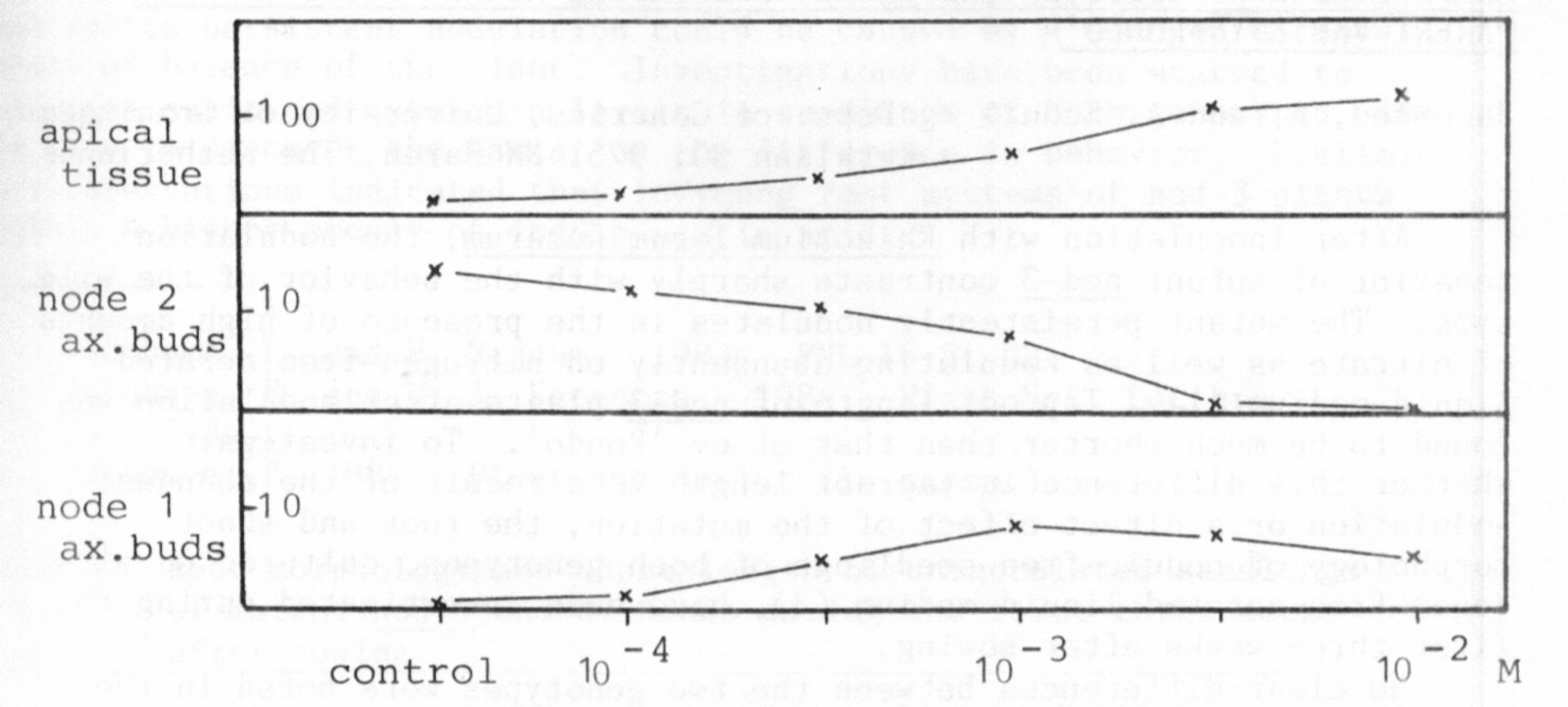
Fig. 1 shows the typical morphogenetic reaction pattern of IL seed-
lings in this assay after apical application of 2,4-1) with increasing
amount. 1) Apical shoot tissue swelled and showed callus structures with
increasing concentrations of 2,4-1) up to 10-2 M. 2) Axillary buds at the
second node were inhibited in elongation and often swollen with in-
creasing amounts of 2,4-D which is in contrast to the control where these
buds elongated and were the dominant regenerating shoots, 3) Axillary
buds at the first node showed a maximum elongation at about 10-3 M, but
were inhibited at higher concentrations. The IL control normally shows no
bud elongation growth at the first node. At the concentration of 10-2 M
2,4-D a characteristic morphogenetic reaction pattern was evident which
is similar to the well known "herbicida1" morphogenetic response.
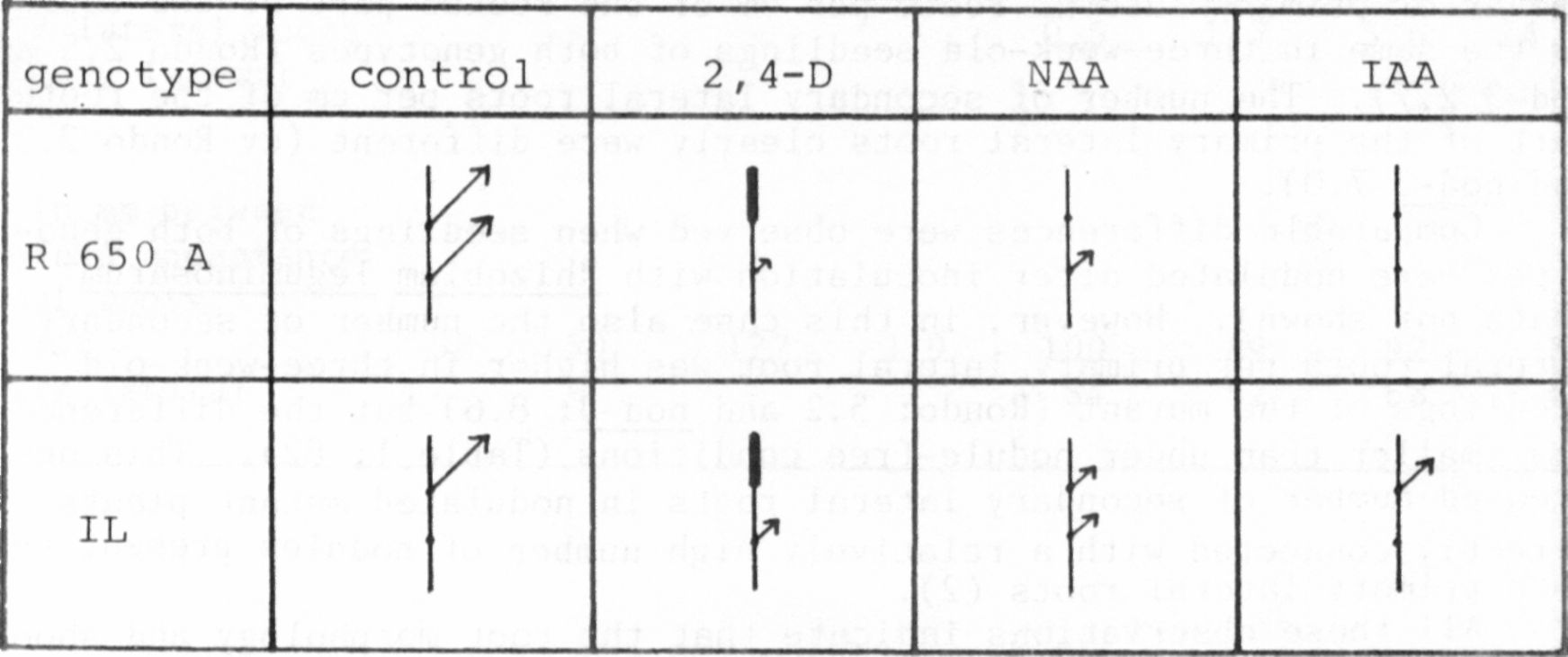
Now, the other auxins, NAA and LAA, were compared to 2,4-D and the
control at these higher concentrations (10-2 M) using seedlings of IL
(Fig. 2, below). 1) The NAA- and IAA-treated seedlings did not show the
drastic tissue proliferation in the apical region that 2,4-D did. 2)
Axillary buds at the second node were more inhibited in elongation com-
pared to the control, but less inhibited compared to the 2,4-D seedlings.
Lowest inhibition was observable after IAA-treatment, 3) Axillary bud
elongation in the first node was inhibited similar to the control in the
case of IAA and a stimulation of bud elongation was observed after NAA—
treatment as in the case of 2,4—D.
1/ This paper is dedicated to Prof. Dr. W. Gottschalk, who kindly sup-
ported my scientific work over the past years at our Institute.