PNL Volume 16 1984 RESEARCH REPORTS
SUPPLEMENTAL LINKAGE DATA FOR CHROMOSOMES 1 AND 5
Marx, G. A. NYS Agricultural Experiment Station, Geneva, NY USA
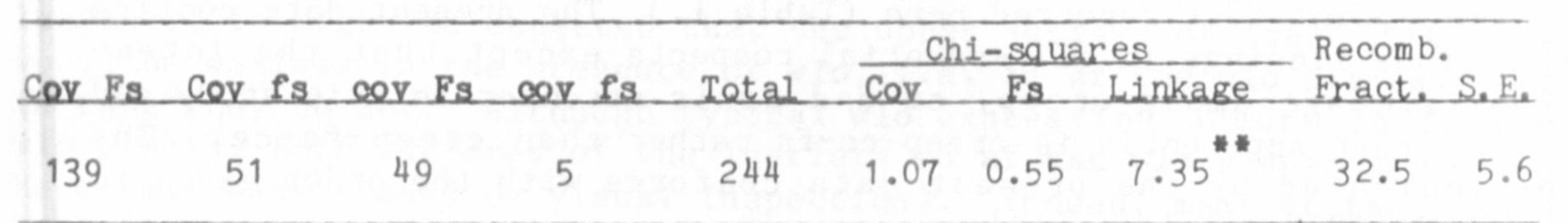
Creep-Ce-Fs
In 1982 (5) I reported evidence showing that creep is situated on
chromosome 5. With respect to other markers on that chromosome, the
following order was suggested: Cr-Creep-Fs-Ce. A line homozygous reces-
sive for creep ce fs was recovered from the above study and was used in
the three-point test reported here (Table 1.) The present data confirm
the previous findings in all essential respects except that the inten-
sity of linkage among creep, ce and fs is stronger in this study and
that the order apparently is creep-ce-fs rather than creep-fs-ce. The
order indicated by the present data conforms with the order given in
Lamprecht's 1968 map [see (1)].
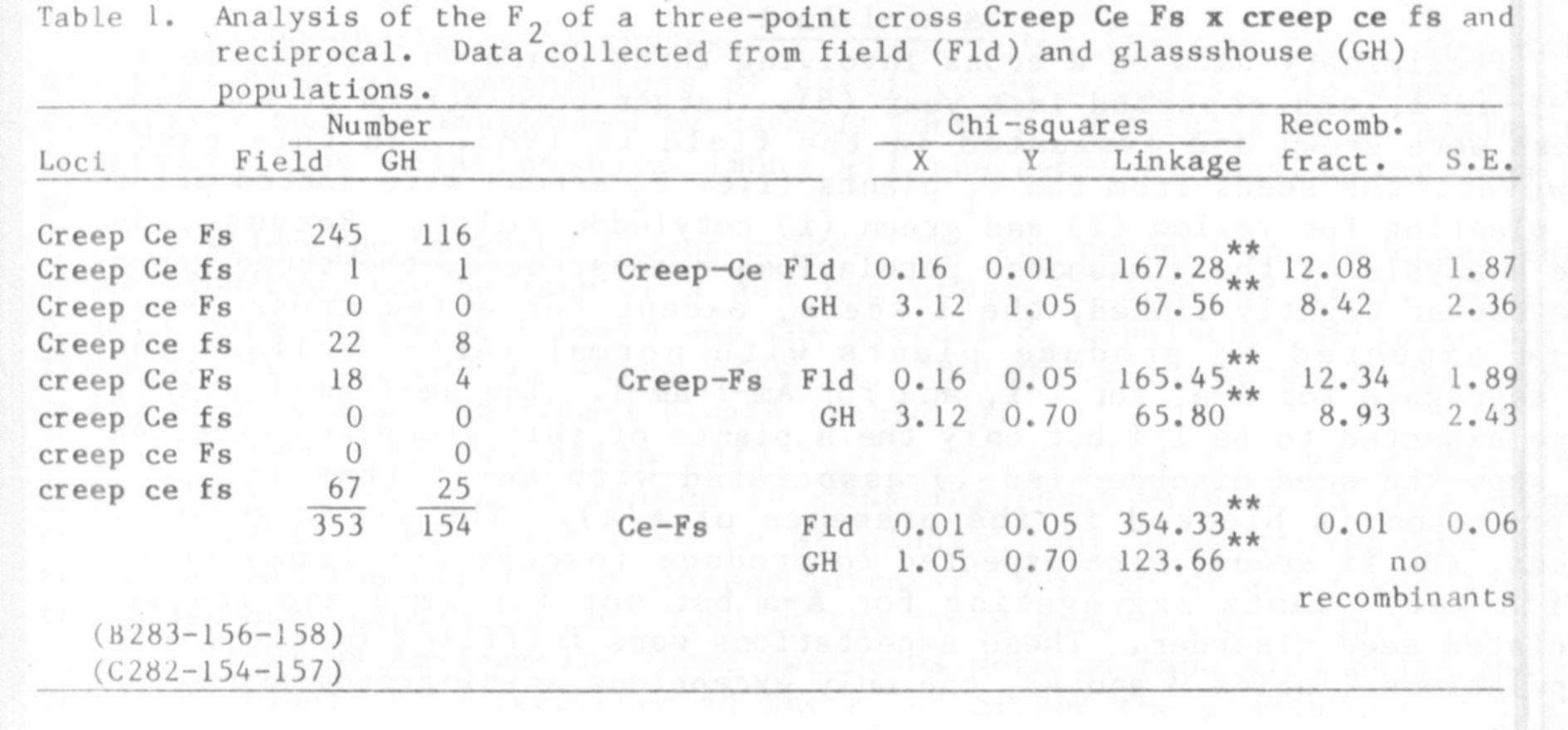
The data are divided into two separate analyses because the overall
experiment was conducted in part in the glasshouse and in part in the
field. There was some question about the genotype of 18 F2 segregants,
so these plants were progeny tested to verify their genotype. There
were only two Ce fs crossover F2 plants in the combined populations
comprising 507 plants.