THE SEGREGATION OF MUTANT GENES
Gottschalk, W. Institute of Genetics, University of Bonn
Federal Republic of Germany
A clear 3:1 segregation in the progenies of monohybrid plants can
be expected only if three conditions are satisfied:
- The frequency of gametes with the dominant and the recessive
allele must be approximately equal.
- The two categories of gametes must have equal chances of par-
ticipating in fertilization.
- The three genotypes AA, Aa, and aa in the F2 or M2 families
must have approximately equal chances of survival during the
earliest stages of ontogenetic development.
The first condition is usually fulfilled because meiosis is a very
reliable process. In the large group of lethal mutants, the third
condition is not fulfilled in all those cases in which the mutant
gene is effective during seed germination or immediately afterwards,
resulting in a deficit of mutants.
In the second and most interesting group, the growth rate of the
pollen tubes containing the mutant recessive allele often is lower than
that of the tubes containing the non-mutant dominant allele. This
phenomenon, called zertation, necessarily leads to a deficit of mutants
in the segregating families, the extent of the deficit depending on the
differences in the growth rates between A and a/a tubes.
This problem cannot be studied in the M2 families because many M1
plants of the garden pea are chimeras composed of non-mutated sectors
and sectors heterozygous for the mutant genes. Segregation can only be
expected in the offspring derived from the heterozygous sectors.
Therefore, the deficit of mutants in the M2 families depends on the size
of the mutated sector of the M1 plants. The chimerical status of the M1
plants, however, influences only the M2 segregations. In later gener-
ations, the "true" segregations can be evaluated.
This problem has been studied using 324 different recessive genes
of the Pisum genome in segregating families of the M3 to M6 generations.
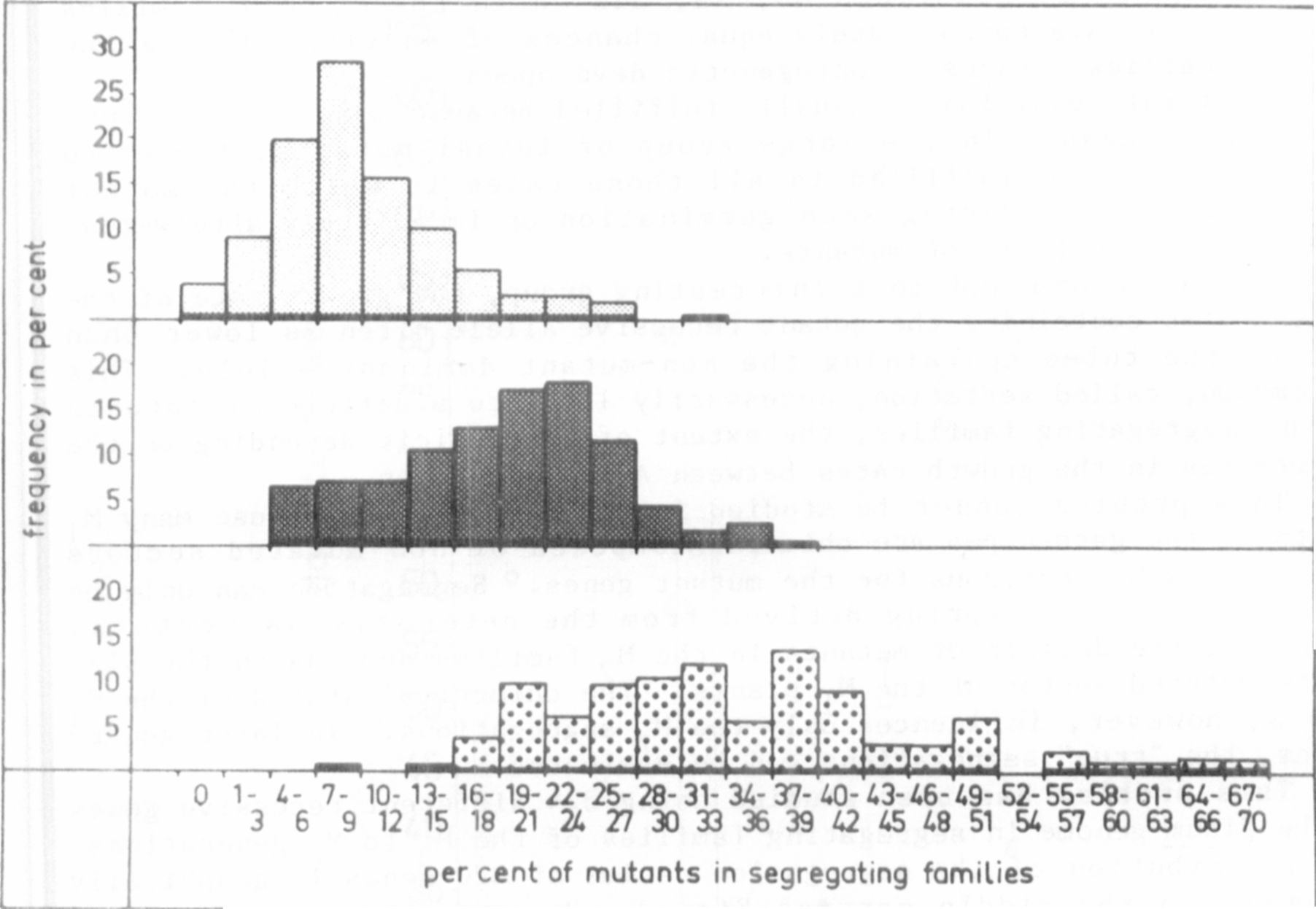
The distribution of the segregation ratios of 320 genes is graphically
presented in the middle part of Fig. 1, showing a clear deficit of
mutants. In many cases, the deviation from a 3:I segregation was
statistically not significant, due to the small number of plants in some
families. If, however, we consider the whole material evaluated, com-
prising 3,523 segregating families, the trend toward a deficit of
mutants becomes clear.
A particularly strong deviation was found in mutants 60A, 168 and
1206A (upper part of Fig. 1). Genotypes 168 and I206A are fertile
chlorophyll mutants; 60A is a tiny, long-living lethal mutant. The
deficit of mutants, it must be emphasized, was not due to an ontogeneti-
cally early death of some of the mutant plants. Similar deficits were
found in subsequent generations, not only In these 3 genotypes, but also
in many mutants belonging to the middle group of the graph.
Furthermore, they were observed in F2 to F4 generations following
hybridizations between the mutants and the mother variety, demonstrating
thereby that these deviations are characteristic peculiarities of the
respective genes.