That the gene Le has a proximal location in the short arm of
chromosome 4 has been mentioned in the paper published in PNL Vol. 12
cited above and this is also strongly supported by the results from my
investigation of a T(4-7)a translocation (4). My next goal has been to
establish the location of a good marker gene viz. N in the long arm of
the same chromosome. L-110 is recessive and L-114 dominant for this
gene and consequently I made a cross between these lines in 1979. In
the F2 of this cross I obtained 189 diploid and 4 trisomic plants, the
latter with an average pollen sterility of 27%. Of the diploid plants
89 were fertile with the ratio 55 N:34n and among the 100 semisterile
diploid plants the ratio was 84N:l6n, the chi-square values being
Fert:Semister. (1:1) = 0.640, N:n = (3:1) 0.213, and T-N = 12.150. The
crossing-over value between T. and N. according to the maximum likelihood
method is 17.8 + 4.50%. However, because of poor germination only 80% of
the seed sown gave rise to plants with mature pods, so some caution must
be observed in interpreting the results. Further investigations would
be desirable using lines concomitantly recessive for several genes of
chromosome 4, e.g. fa, was, and n. instead of L-110.
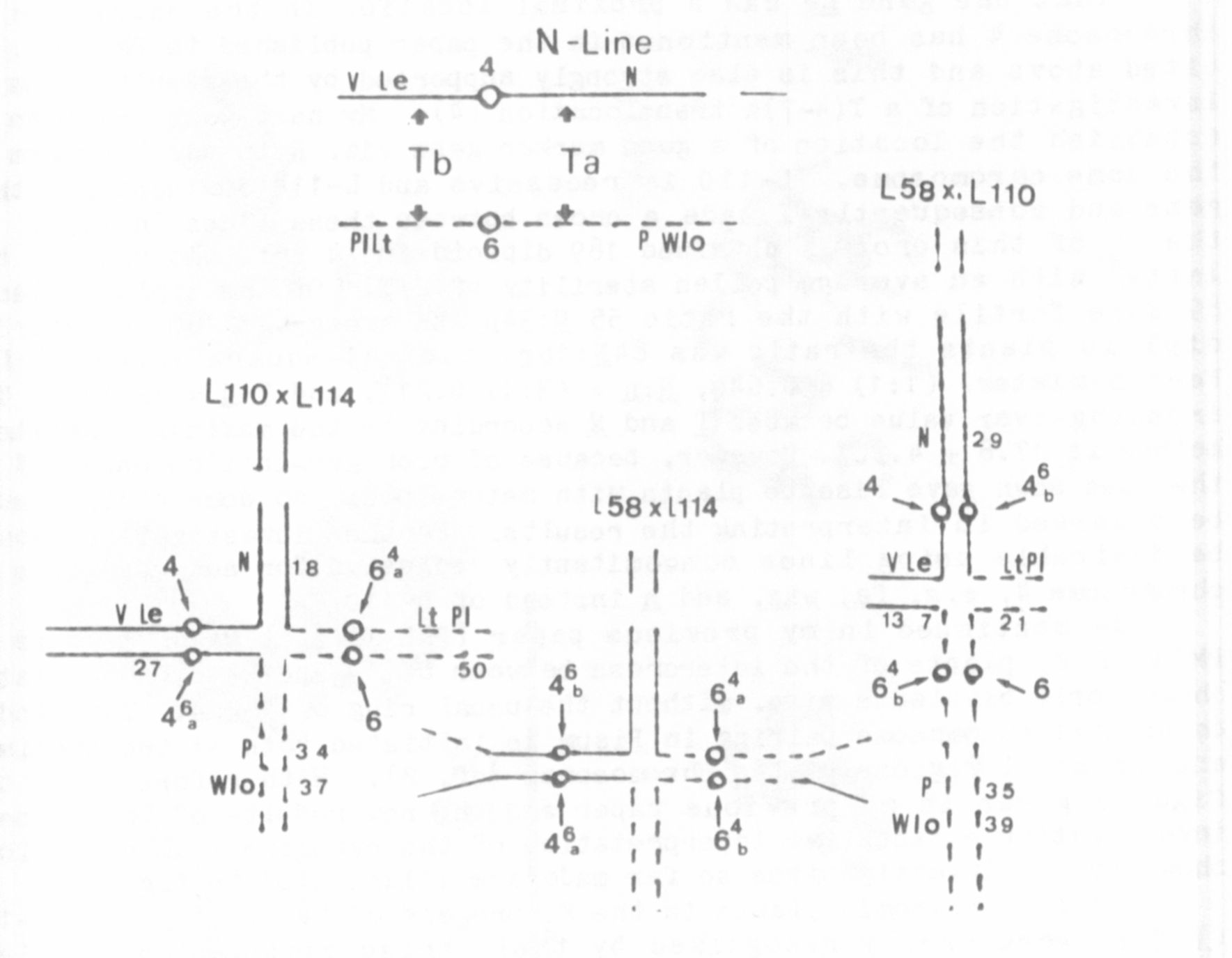
As mentioned in my previous paper (PNL 12), at MI of meiosis in
PMCs of F plants of the intercross between L-114 and L-58, some plates
showed only bivalents (j.e. without the usual ring of four). This indi-
cates that chromosome pciring in Pisum is initiated both at the proximal
and terminal regions of the chromosomes (cf. 2). With reference to the
figures given in my previous paper and the new results of the present
investigation a tentative interpretation of the cytogenetical conditions
shown by the investigations so far made are illustrated in Fig. 2.
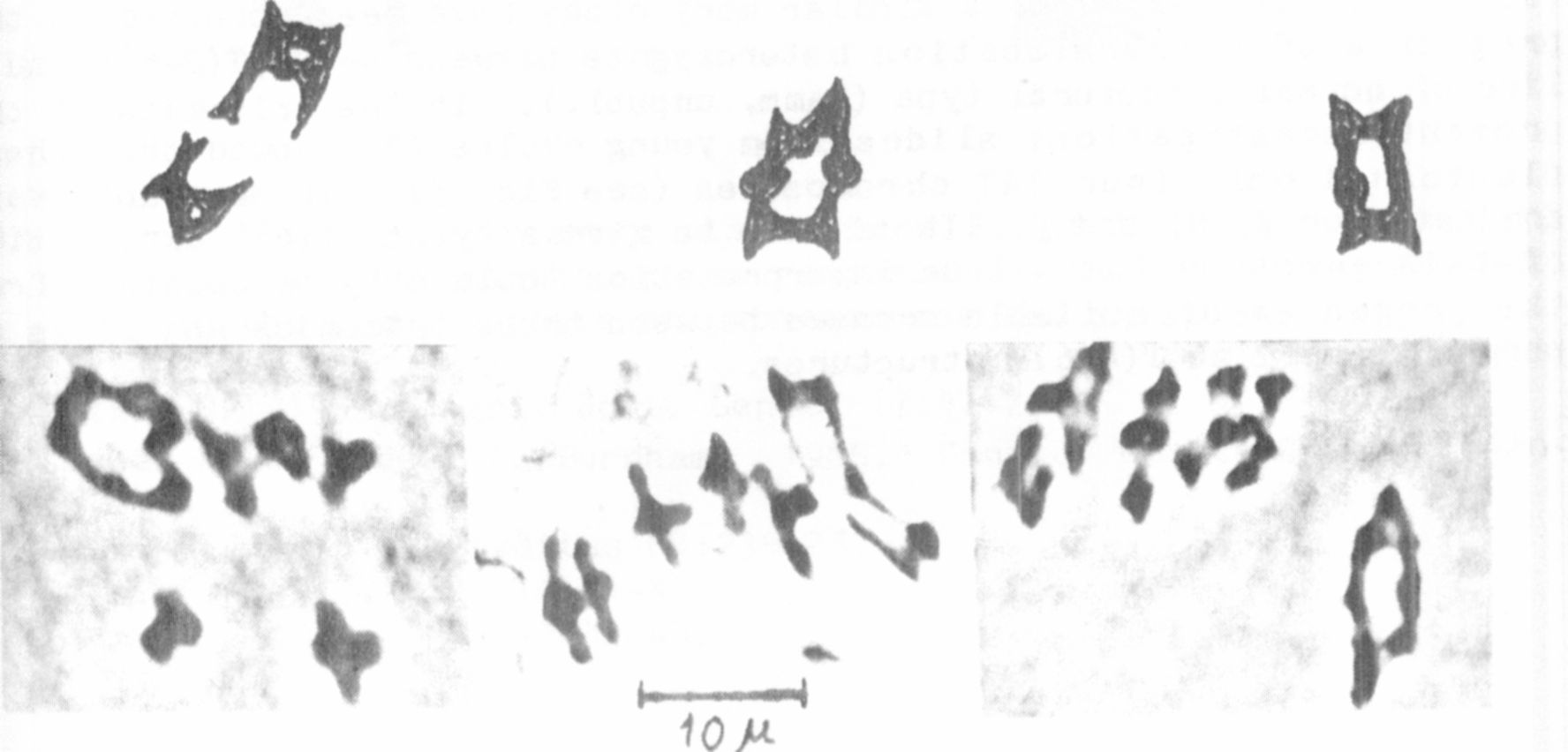
The four trisomic plants in the F progeny of L-110 (.y. n p) x L-114
(V N P) were easily recognized by their broad stipules and leaflets
(Fig. 3). Trisomies of a similar morphology have been observed in the
progenies of a translocation heterozygote between L-108,T(2-6)a and a
line of normal structural type (Lamm, unpubl.). In the trisomies of the
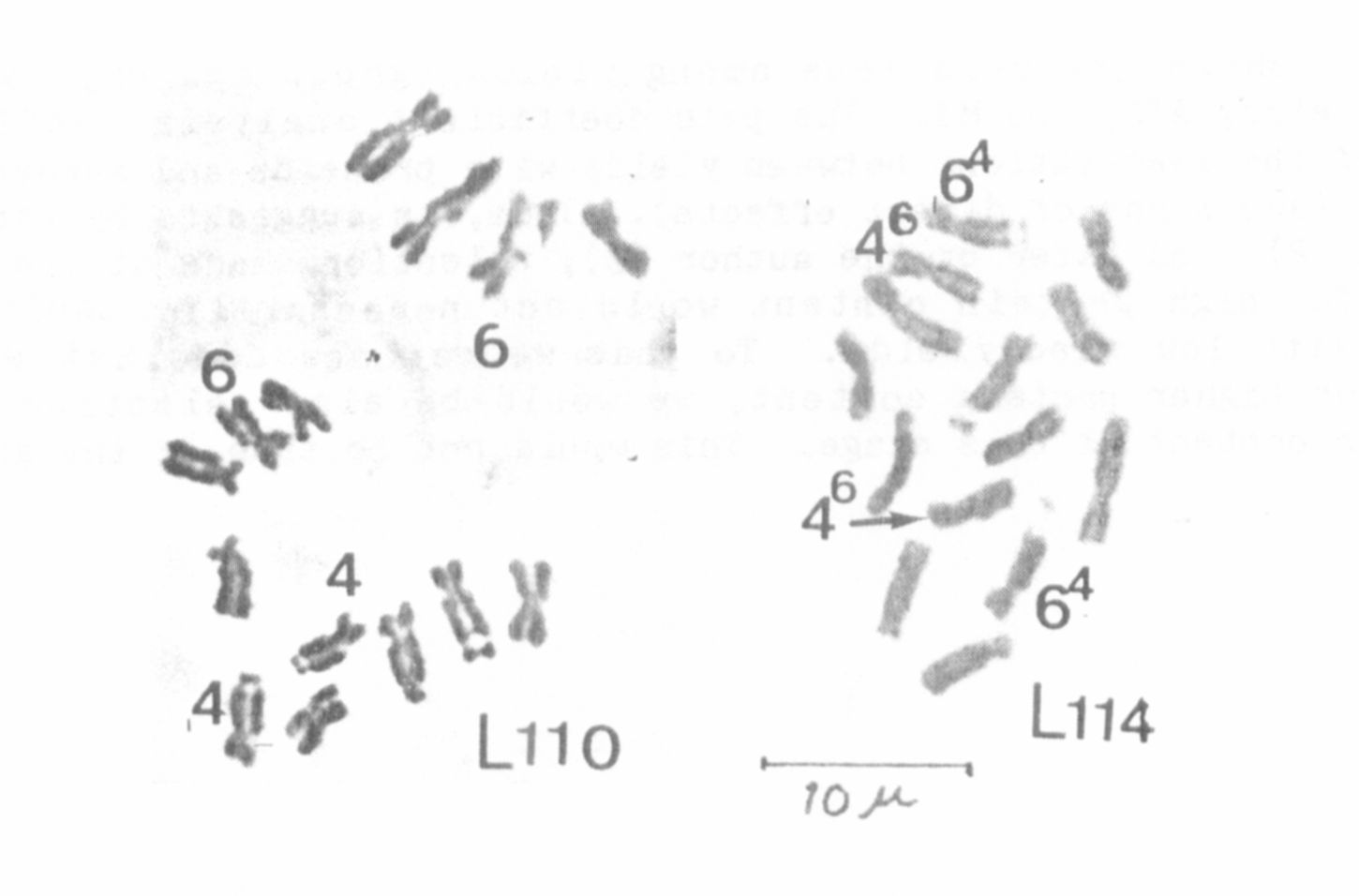
present investigations slides from young ovules (3) showed that these
plants had only four SAT chromosomes (see Fig. 3). These plants were
dominant for V, N and P. It is easy to give a hypothetical explanation
of this genotype but a true interpretation could only be obtained from
the progenies of suitable crosses between these trisomies and lines of
normal as well as T(4-6)a structures.