44 RESEARCH REPORTS
PNL Volume 12 1980
FASCIATION AND HETEROSIS IN PEA
Lonnig, W. E.
Institute of Genetics, Bonn, Federal Republic of Germany
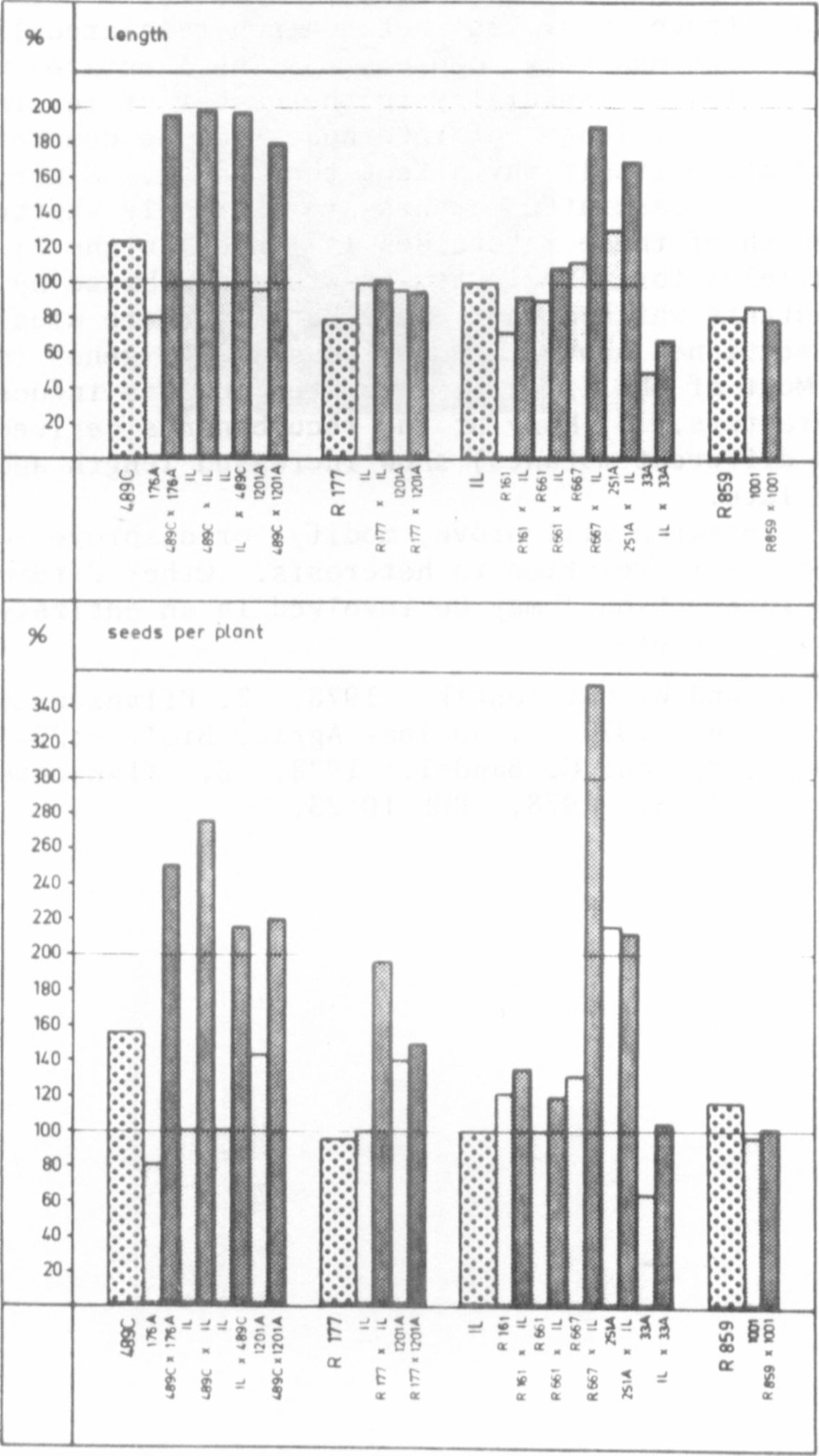
Under the climatic conditions of Central Europe, our fasciated mutant
489 C always causes a strong heterotic effect in stem length, seeds per plant,
and other characters when crossed with the initial line (IL) 'Dippes gelbe
Viktoria', other lines, or different mutants (Fig. 1). Mutants 251 A, 123,
and other fasciated forms genetically comparable with mutant 489 C cause a
similar, but in some cases clearly weaker, heterotic effect (2).
As perhaps more than 15 genes are mutated in
489 C, it is not possible to say prima facie whether

one or more of the genes causing fasciation or other
unrelated mutant genes contribute to the heterotic
effects. Nor is it clear whether the phenomenon
is caused by a) heterozygosity per se, b) dominant
factors "hidden" or impeded by detrimental recessive
genes in the same mutant [most of the 15 genes have
mutated to the recessive state, as, for instance,
the genes for fasciation, but a few of them are
dominant in relation to the IL, as lateness in
flowering and ripening, sensitivity to day length
(4), and, perhaps, the genes for increased stem
length], c) gene interactions, or d) a combination
of the above.
In seeking answers to these questions, we used
the following mutants and recombinants for our
studies (all forms are derived from Dippes gelbe
Viktoria): a) strongly fasciated mutants 489 C,
33 A, b) linearly fasciated mutant 251 A, c) linearly
fasciated recombinants R 859 (derived from 123 x
46 C), R 661 (derived from 489 C x 26), and R 667
(derived from 489 C x 169), d) weakly fasciated
recombinants, stem bifurcated, R 161 (derived from
489 C x 1201 A) and R 177 (derived from 489 C x
1202 A), e) other mutants 176 A—narrow leaves,
flowers, and pods, 1201 A—stem bifurcated, 1001 —
Fig. 1.
489C 1201A
489C
x
1201A
thousand-grain weight increased.
The results of these crosses are shown in Fig. 2
and can be summarized as follows: a) 33 A, R 161,
R 661, and R 859 do not cause heterosis; the small
deviations are well in the range of the IL. This
means that the genetic causes for strong, linear, and weak fasciation are
not necessarily correlated with the heterotic effects, b) R 177 x IL, etc.,
do not show heterosis in length although the character "seeds per plant" seems
to show significant heterosis. For final analysis the crosses should be
repeated with more extensive material. If verified, there would be no strict
correlation between heterosis in length and yield, c) linearly fasciated
R 667 seems to cause a strong heterotic effect, comparable to that of 489 C
x IL, etc. However, 667 x IL is not very representative as we had only nine