PNL Volume 12 1980
RESEARCH REPORTS 43
L-114 and L-58 have been crossed with Klein's T-488, which is homozygous
for two reciprocal translocations, viz. T(2-3)a and T(4-7)a (cf. R. Lamm,
PNL 10:31-32, 1978). The observed maximum configurations at M I of meiosis
given in Table 1 are in agreement with those expected. Whereas the maximum
configuration of one ring of six chromosomes + one ring of four + two bivalents
is very common in the F1 of cross No. 5 of Table 1, it is rare in cross No.
6 where it may easily be overlooked. In this F1 hybrid, configurations
with two rings of four and three bivalents dominate. From this point of
view, L-114 is superior to L-58 as a member of a translocation tester set.
On the other hand, the linkage relations between the T-points of L-58 and
marker genes in the chromosomes involved are, so far, better known in this
line than in L-114. The gametic sterility is equal for the two F1 hybrids
of the crosses Nos. 5 and 6, both with respect to the male and female gametes,
which is remarkable considering the differential pairing behavior.
In H. Lamprecht's "Monographic der Gattung Pisum" (Graz, 1974) linkage
values between the T-points of L-58 and the genes Le, N, Wlo, P, and Lt
are given on p. 492. Especially the two genes first mentioned show strong
linkage with the T-point. These genes are certainly situated in the short
arm of chromosome No. 4. In the investigations of R. Lamm and R. J. Miravalle
(Hereditas 45:417-440, 1959), there was no significant linkage between the
T-points of L-114 and the genes Le and PI. These data have been completed
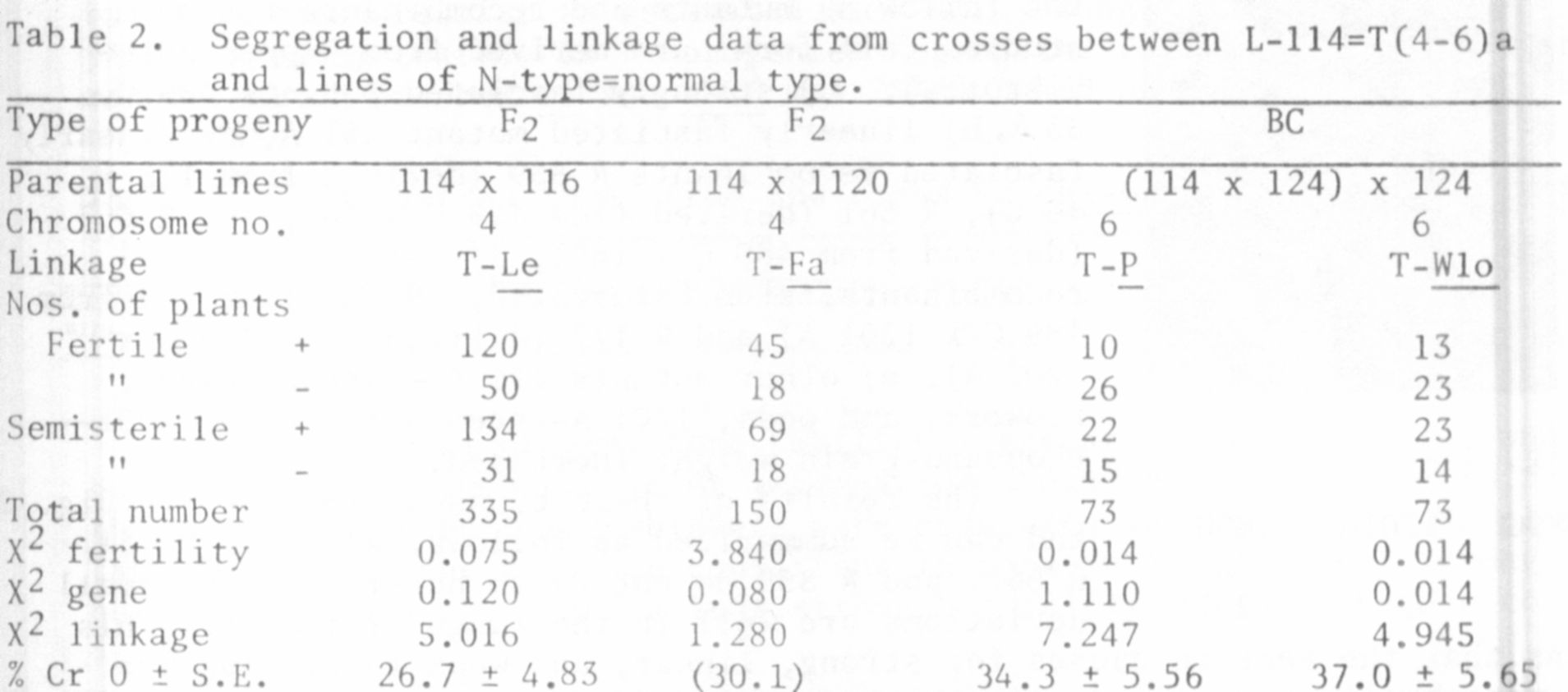
by the results given in Table 2, which indicate linkage relations to the
genes Le and Fa of chromosome No. 4 and P and Wlo of chromosome No. 6. The
backcross data of Table 2 originate from investigations made by Ernst Nilsson
(unpub.). The probable linkage between the T-points and the gene N of chromo-
some No. 4 will be tested by investigations in progress.
As regards the intercross method, future work on this subject would be
greatly facilitated by trying to find translocation lines involving the same
two chromosomes but with the T-points situated in the same arm of one of the
chromosomes but in different arms of the other chromosome. This is especially
desirable for the mapping work of chromosomes 4 and 7 since, in my experience,
translocation heterozygotes involving these chromosomes seldom, if ever, give
rise to interchange trisomies in their progenies. Such trisomies are otherwise
one of the best tools for the mapping of the chromosomes.